

A dual role for the histone methyltransferase PR-SET7/SETD8 and histone H4 lysine 20 monomethylation in the local regulation of RNA polymerase II pausing
- PMID: 24459145
- PMCID: PMC3953257
- DOI: 10.1074/jbc.M113.520783
A dual role for the histone methyltransferase PR-SET7/SETD8 and histone H4 lysine 20 monomethylation in the local regulation of RNA polymerase II pausing
Abstract
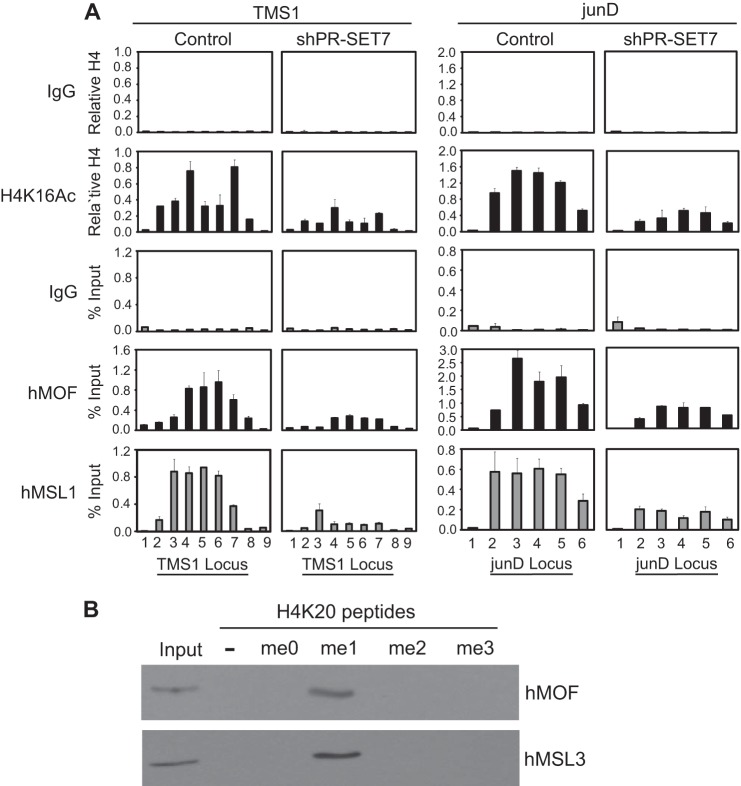
RNA polymerase II (Pol II) promoter-proximal pausing plays a critical role in postinitiation transcriptional regulation at many metazoan genes. We showed recently that histone H4 lysine 16 acetylation (H4K16Ac), mediated by the MSL complex, facilitates the release of paused Pol II. In contrast, H4 lysine 20 trimethylation (H4K20me3), mediated by SUV420H2, enforces Pol II pausing by inhibiting MSL recruitment. However, how the balance between H4K16Ac and H4K20me3 is locally regulated remains unclear. Here, we demonstrate that PR-SET7/SETD8, which monomethylates histone H4 lysine 20 (H4K20me1), controls both H4K16Ac and H4K20me3 and in doing so, regulates Pol II pausing dynamics. We find that PR-SET7-mediated H4K20me1 is necessary for the recruitment of the MSL complex, subsequent H4K16Ac, and release of Pol II into active elongation. Although dispensable for SUV420H2 recruitment, PR-SET7-mediated H4K20me1 is required for H4K20me3. Although depletion of SUV420H2 is sufficient to deplete H4K20me3 and relieve an H4K20me3-induced pause, pausing is maintained in the absence of PR-SET7 despite H4K20me3 depletion because of an inability to recruit the MSL complex in the absence of H4K20me1. These findings highlight the requirement for PR-SET7 and H4K20me1 in establishing both the H4K16Ac and H4K20me3 marks and point to a dual role in the local regulation of Pol II pausing.
Keywords: Chromatin Histone Modification; Chromatin Structure; Gene Regulation; H4K20me1; MSL Complex; PR-SET7/SETD8; RNA Polymerase; RNA Polymerase II Promoter-proximal Pausing; SUV420H2; Transcription Regulation.
Figures





References
-
- Yamaguchi Y., Takagi T., Wada T., Yano K., Furuya A., Sugimoto S., Hasegawa J., Handa H. (1999) NELF, a multisubunit complex containing RD, cooperates with DSIF to repress RNA polymerase II elongation. Cell 97, 41–51 - PubMed
-
- Cheng B., Price D. H. (2007) Properties of RNA polymerase II elongation complexes before and after the P-TEFb-mediated transition into productive elongation. J. Biol. Chem. 282, 21901–21912 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
