

Targeted gene mutation of E2F1 evokes age-dependent synaptic disruption and behavioral deficits
- PMID: 24460902
- PMCID: PMC4028376
- DOI: 10.1111/jnc.12655
Targeted gene mutation of E2F1 evokes age-dependent synaptic disruption and behavioral deficits
Abstract
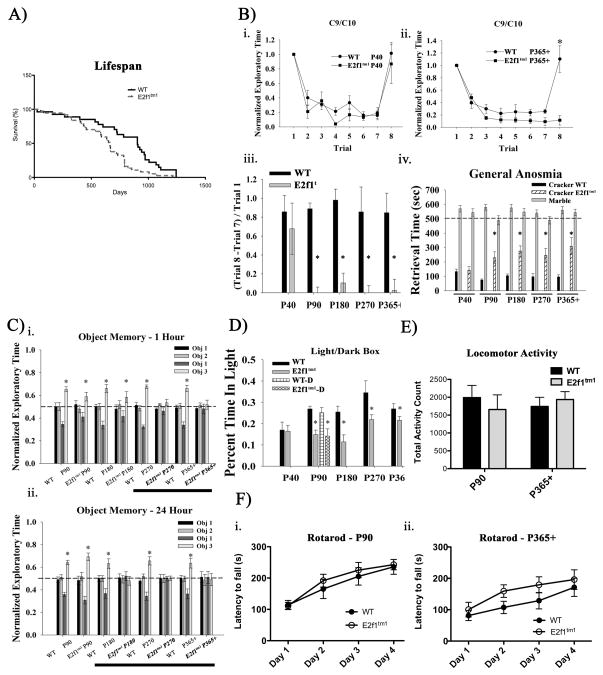
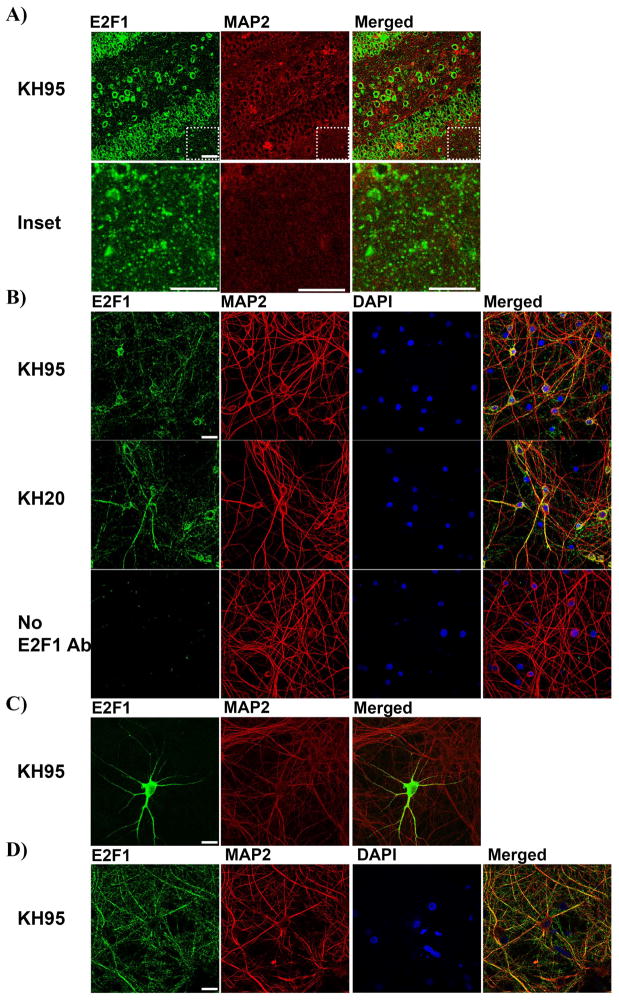
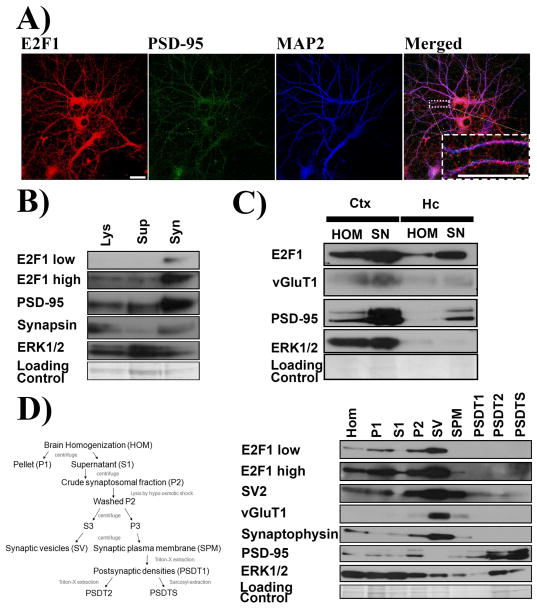
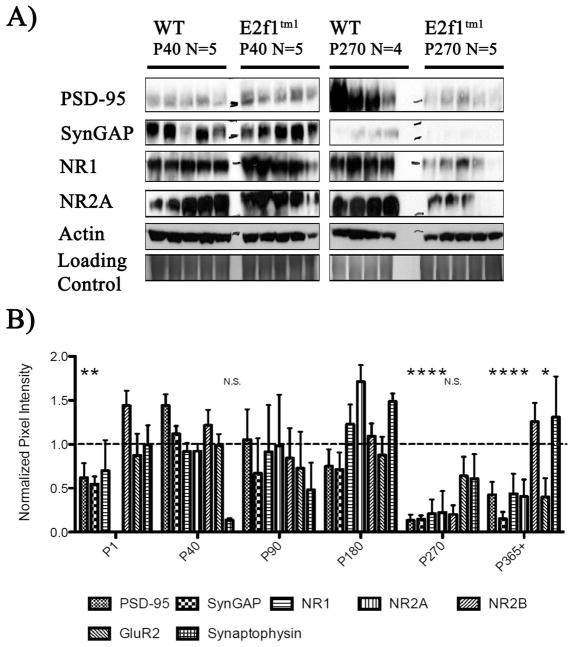
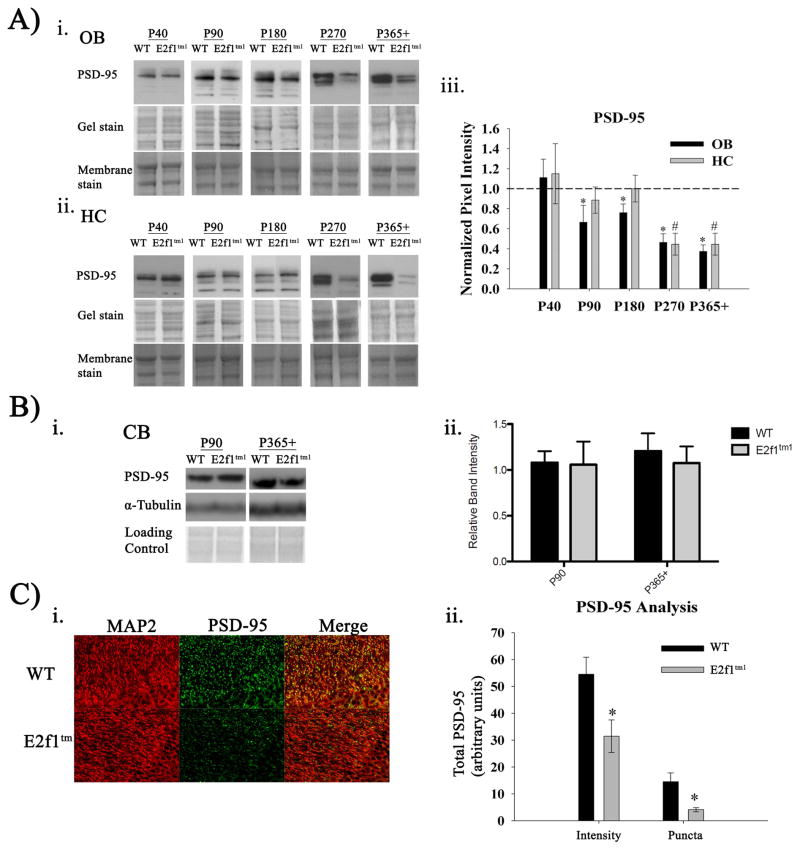
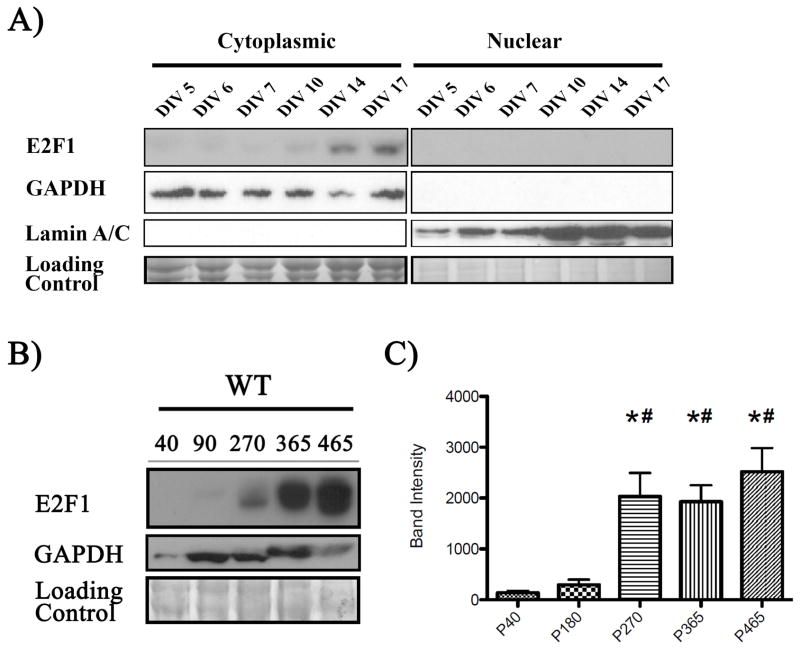
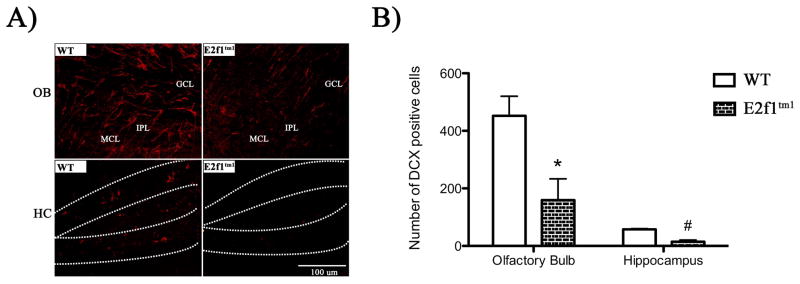
Aberrant expression and activation of the cell cycle protein E2F1 in neurons has been implicated in many neurodegenerative diseases. As a transcription factor regulating G1 to S phase progression in proliferative cells, E2F1 is often up-regulated and activated in models of neuronal death. However, despite its well-studied functions in neuronal death, little is known regarding the role of E2F1 in the mature brain. In this study, we used a combined approach to study the effect of E2F1 gene disruption on mouse behavior and brain biochemistry. We identified significant age-dependent olfactory and memory-related deficits in E2f1 mutant mice. In addition, we found that E2F1 exhibits punctated staining and localizes closely to the synapse. Furthermore, we found a mirroring age-dependent loss of post-synaptic protein-95 in the hippocampus and olfactory bulb as well as a global loss of several other synaptic proteins. Coincidently, E2F1 expression is significantly elevated at the ages, in which behavioral and synaptic perturbations were observed. Finally, we show that deficits in adult neurogenesis persist late in aged E2f1 mutant mice which may partially contribute to the behavior phenotypes. Taken together, our data suggest that the disruption of E2F1 function leads to specific age-dependent behavioral deficits and synaptic perturbations. E2F1 is a transcription factor regulating cell cycle progression and apoptosis. Although E2F1 dysregulation under toxic conditions can lead to neuronal death, little is known about its physiologic activity in the healthy brain. Here, we report significant age-dependent olfactory and memory deficits in mice with dysfunctional E2F1. Coincident with these behavioral changes, we also found age-matched synaptic disruption and persisting reduction in adult neurogenesis. Our study demonstrates that E2F1 contributes to physiologic brain structure and function.
Keywords: E2F; adult neurogenesis; cell cycle; synaptic proteins; transgenic mice.
© 2014 International Society for Neurochemistry.
Conflict of interest statement
Conflicts of interest:
“The authors have no conflict of interest to declare.”
Figures
References
-
- Barresi M, Ciurleo R, Giacoppo S, Foti Cuzzola V, Celi D, Bramanti P, Marino S. Evaluation of olfactory dysfunction in neurodegenerative diseases. Journal of the neurological sciences. 2012;323:16–24. - PubMed
-
- Barrett LE, Sul JY, Takano H, Van Bockstaele EJ, Haydon PG, Eberwine JH. Region-directed phototransfection reveals the functional significance of a dendritically synthesized transcription factor. Nature methods. 2006;3:455–460. - PubMed
-
- Cooperkuhn C, Vroemen M, Brown J, Ye H, Thompson M, Winkler J, Kuhn H. Impaired Adult Neurogenesis in Mice Lacking the Transcription Factor E2F1. Molecular and Cellular Neuroscience. 2002;21:312–323. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
