

Genome-scale reconstruction of the sigma factor network in Escherichia coli: topology and functional states
- PMID: 24461193
- PMCID: PMC3923258
- DOI: 10.1186/1741-7007-12-4
Genome-scale reconstruction of the sigma factor network in Escherichia coli: topology and functional states
Abstract
Background: At the beginning of the transcription process, the RNA polymerase (RNAP) core enzyme requires a σ-factor to recognize the genomic location at which the process initiates. Although the crucial role of σ-factors has long been appreciated and characterized for many individual promoters, we do not yet have a genome-scale assessment of their function.
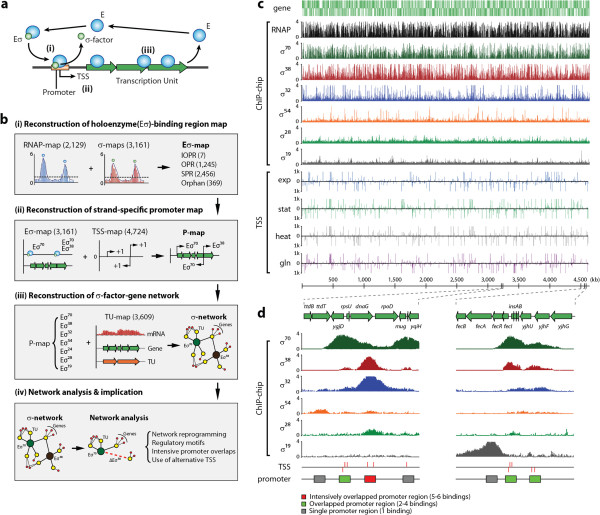
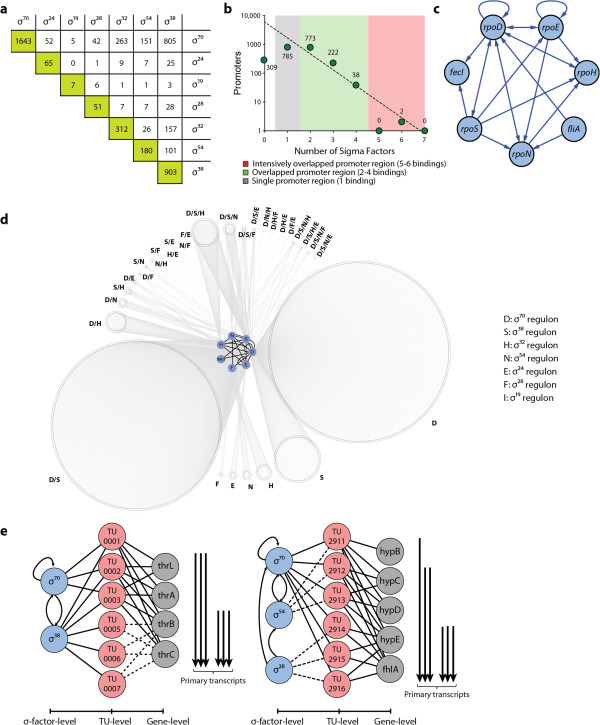
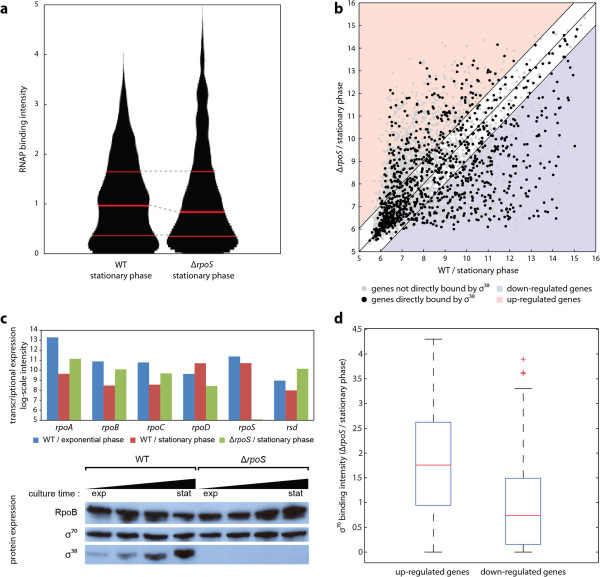
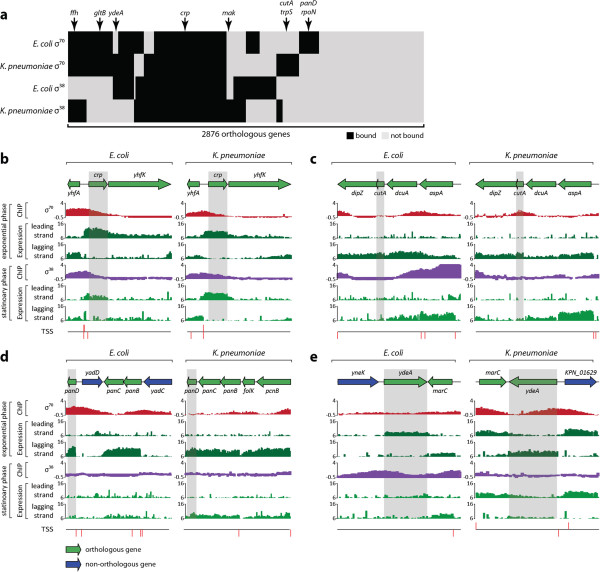
Results: Using multiple genome-scale measurements, we elucidated the network of σ-factor and promoter interactions in Escherichia coli. The reconstructed network includes 4,724 σ-factor-specific promoters corresponding to transcription units (TUs), representing an increase of more than 300% over what has been previously reported. The reconstructed network was used to investigate competition between alternative σ-factors (the σ70 and σ38 regulons), confirming the competition model of σ substitution and negative regulation by alternative σ-factors. Comparison with σ-factor binding in Klebsiella pneumoniae showed that transcriptional regulation of conserved genes in closely related species is unexpectedly divergent.
Conclusions: The reconstructed network reveals the regulatory complexity of the promoter architecture in prokaryotic genomes, and opens a path to the direct determination of the systems biology of their transcriptional regulatory networks.
Figures
Similar articles
-
Isolation and characterization of mutations in region 1.2 of Escherichia coli sigma70.Mol Microbiol. 2001 Oct;42(2):427-37. doi: 10.1046/j.1365-2958.2001.02642.x. Mol Microbiol. 2001. PMID: 11703665
-
The interaction between sigma70 and the beta-flap of Escherichia coli RNA polymerase inhibits extension of nascent RNA during early elongation.Proc Natl Acad Sci U S A. 2005 Mar 22;102(12):4488-93. doi: 10.1073/pnas.0409850102. Epub 2005 Mar 10. Proc Natl Acad Sci U S A. 2005. PMID: 15761057 Free PMC article.
-
Interaction of Escherichia coli RNA polymerase σ70 subunit with promoter elements in the context of free σ70, RNA polymerase holoenzyme, and the β'-σ70 complex.J Biol Chem. 2011 Jan 7;286(1):270-9. doi: 10.1074/jbc.M110.174102. Epub 2010 Oct 15. J Biol Chem. 2011. PMID: 20952386 Free PMC article.
-
The molecular basis of selective promoter activation by the sigmaS subunit of RNA polymerase.Mol Microbiol. 2007 Mar;63(5):1296-306. doi: 10.1111/j.1365-2958.2007.05601.x. Mol Microbiol. 2007. PMID: 17302812 Review.
-
σ70 and PhoB activator: getting a better grip.Transcription. 2012 Jul-Aug;3(4):160-4. doi: 10.4161/trns.20444. Epub 2012 Jul 1. Transcription. 2012. PMID: 22771992 Free PMC article. Review.
Cited by
-
Exploring the role of sigma factor gene expression on production by Corynebacterium glutamicum: sigma factor H and FMN as example.Front Microbiol. 2015 Jul 22;6:740. doi: 10.3389/fmicb.2015.00740. eCollection 2015. Front Microbiol. 2015. PMID: 26257719 Free PMC article.
-
Identification of a transcription factor, PunR, that regulates the purine and purine nucleoside transporter punC in E. coli.Commun Biol. 2021 Aug 19;4(1):991. doi: 10.1038/s42003-021-02516-0. Commun Biol. 2021. PMID: 34413462 Free PMC article.
-
Discovery of numerous novel small genes in the intergenic regions of the Escherichia coli O157:H7 Sakai genome.PLoS One. 2017 Sep 13;12(9):e0184119. doi: 10.1371/journal.pone.0184119. eCollection 2017. PLoS One. 2017. PMID: 28902868 Free PMC article.
-
The molecular basis for control of ETEC enterotoxin expression in response to environment and host.PLoS Pathog. 2015 Jan 8;11(1):e1004605. doi: 10.1371/journal.ppat.1004605. eCollection 2015 Jan. PLoS Pathog. 2015. PMID: 25569153 Free PMC article.
-
BosR and PlzA reciprocally regulate RpoS function to sustain Borrelia burgdorferi in ticks and mammals.J Clin Invest. 2023 Mar 1;133(5):e166710. doi: 10.1172/JCI166710. J Clin Invest. 2023. PMID: 36649080 Free PMC article.
References
-
- Sharma UK, Chatterji D. Transcriptional switching in Escherichia coli during stress and starvation by modulation of sigma activity. FEMS Microbiol Rev. 2010;12:646–657. - PubMed
-
- Yamamoto K, Hirao K, Oshima T, Aiba H, Utsumi R, Ishihama A. Functional characterization in vitro of all two-component signal transduction systems from Escherichia coli. J Biol Chem. 2005;12:1448–1456. - PubMed
-
- Herring CD, Raffaelle M, Allen TE, Kanin EI, Landick R, Ansari AZ, Palsson BO. Immobilization of Escherichia coli RNA polymerase and location of binding sites by use of chromatin immunoprecipitation and microarrays. J Bacteriol. 2005;12:6166–6174. doi: 10.1128/JB.187.17.6166-6174.2005. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
