

Interplay between pre-mRNA splicing and microRNA biogenesis within the supraspliceosome
- PMID: 24464992
- PMCID: PMC3985634
- DOI: 10.1093/nar/gkt1413
Interplay between pre-mRNA splicing and microRNA biogenesis within the supraspliceosome
Abstract
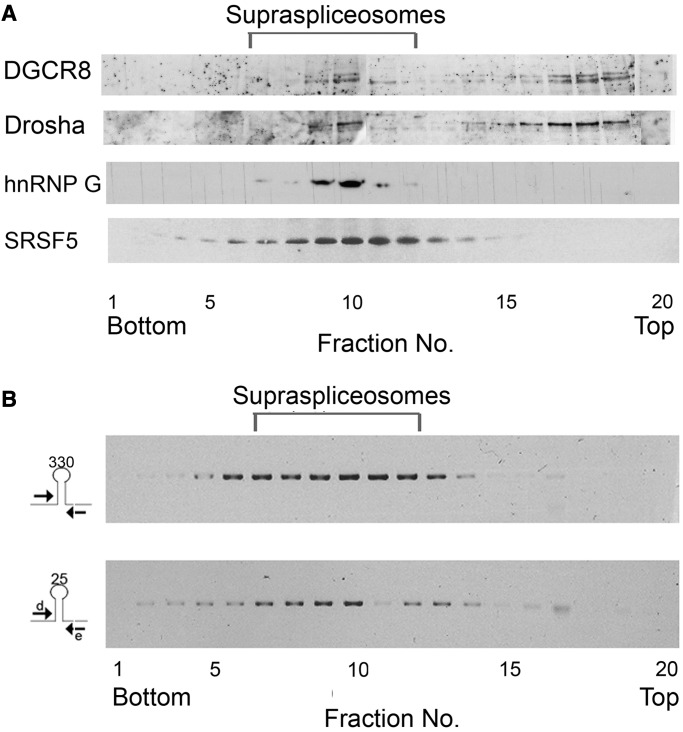
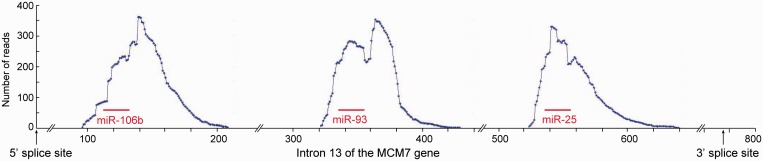
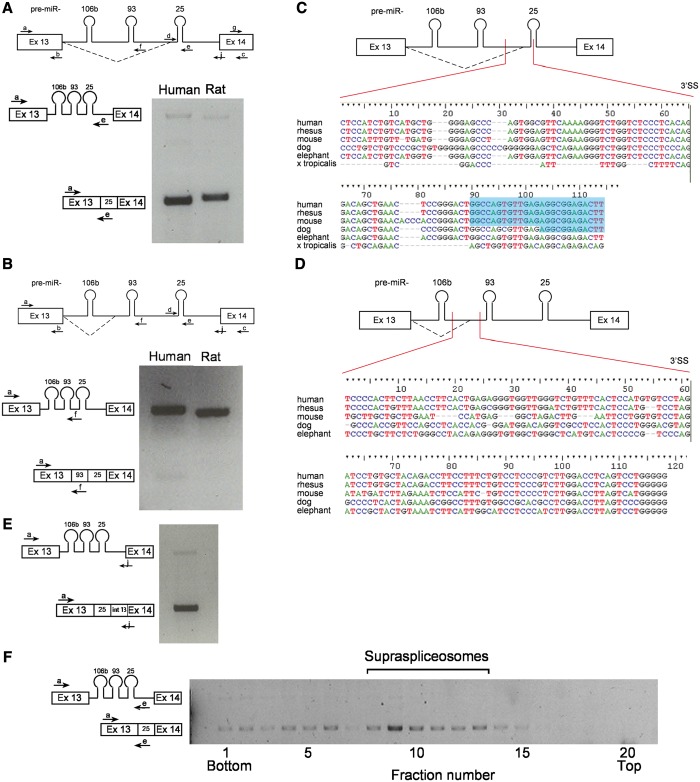
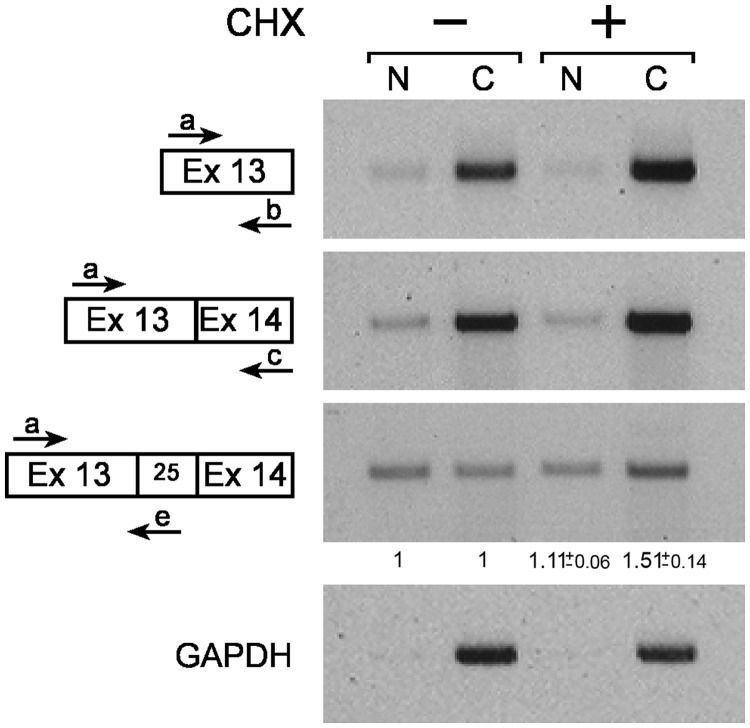
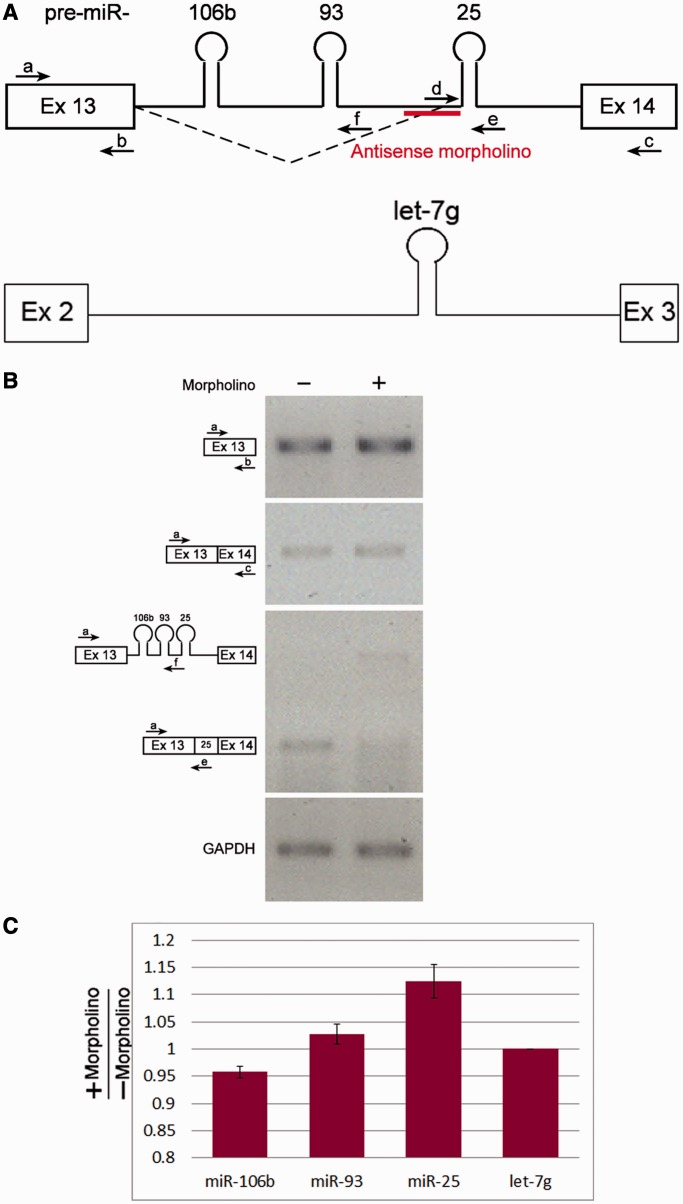
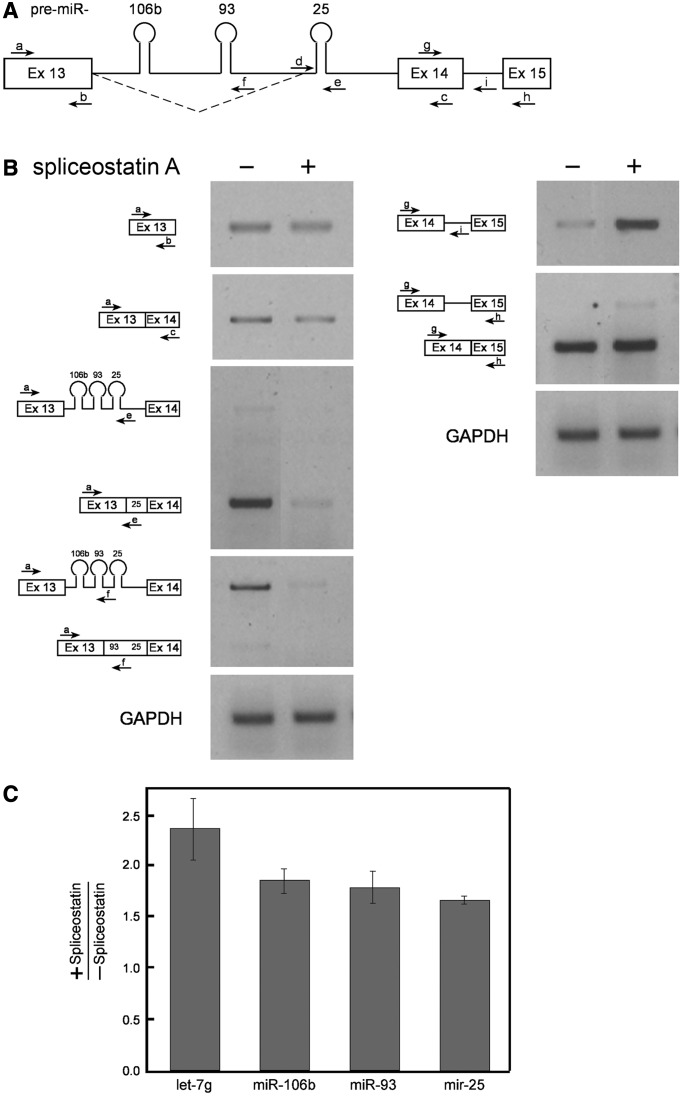
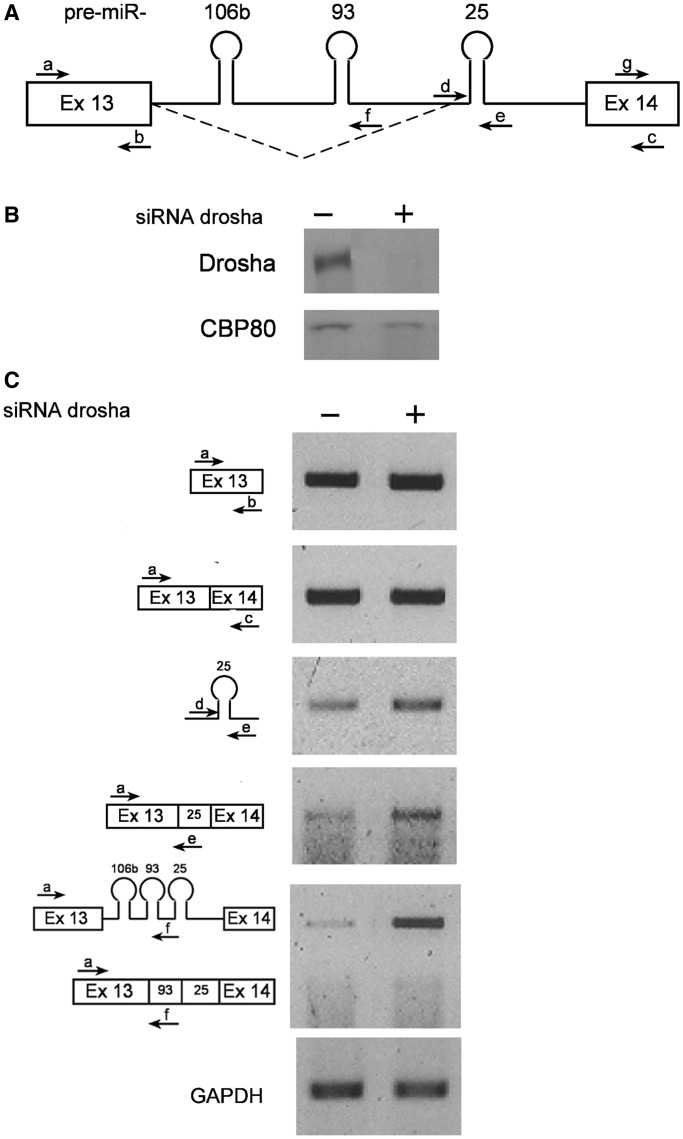
MicroRNAs (miRNAs) are central regulators of gene expression, and a large fraction of them are encoded in introns of RNA polymerase II transcripts. Thus, the biogenesis of intronic miRNAs by the microprocessor and the splicing of their host introns by the spliceosome require coordination between these processing events. This cross-talk is addressed here. We show that key microprocessor proteins Drosha and DGCR8 as well as pre-miRNAs cosediment with supraspliceosomes, where nuclear posttranscriptional processing is executed. We further show that inhibition of splicing increases miRNAs expression, whereas knock-down of Drosha increases splicing. We identified a novel splicing event in intron 13 of MCM7, where the miR-106b-25 cluster is located. The unique splice isoform includes a hosted pre-miRNA in the extended exon and excludes its processing. This indicates a possible mechanism of altering the levels of different miRNAs originating from the same transcript. Altogether, our study indicates interplay between the splicing and microprocessor machineries within a supraspliceosome context.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
