

Down-regulation of Rad51 activity during meiosis in yeast prevents competition with Dmc1 for repair of double-strand breaks
- PMID: 24465215
- PMCID: PMC3900393
- DOI: 10.1371/journal.pgen.1004005
Down-regulation of Rad51 activity during meiosis in yeast prevents competition with Dmc1 for repair of double-strand breaks
Abstract
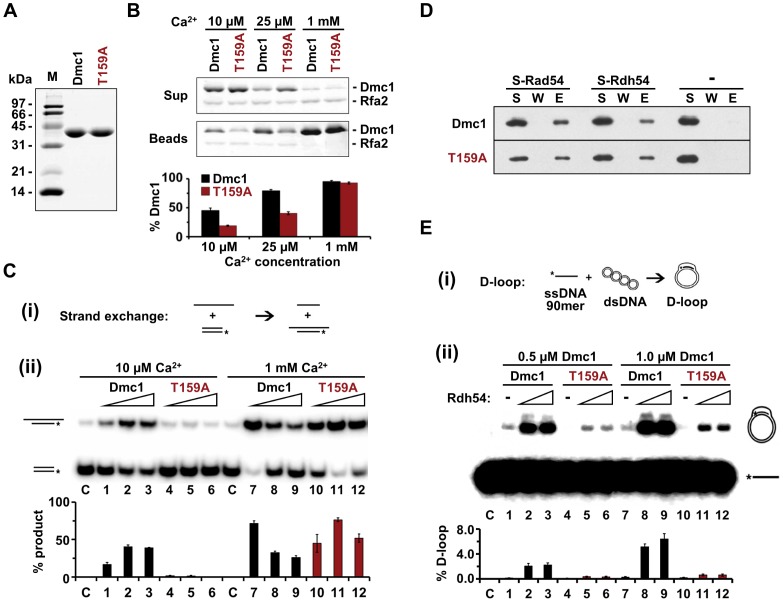
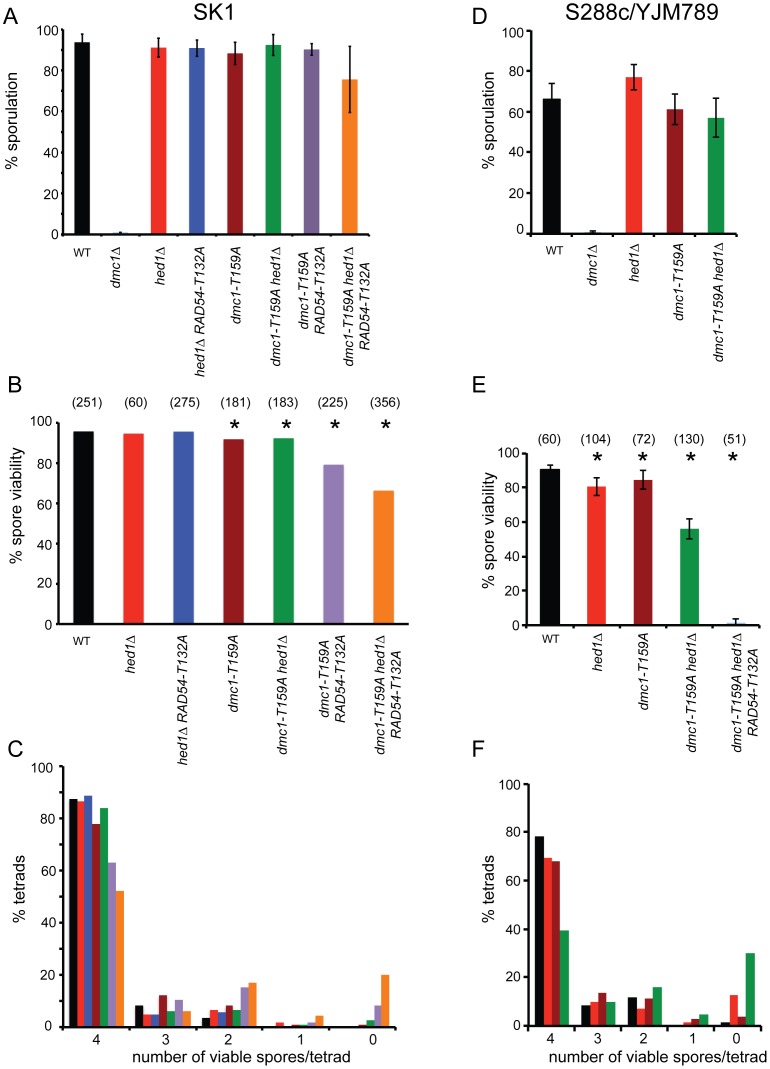
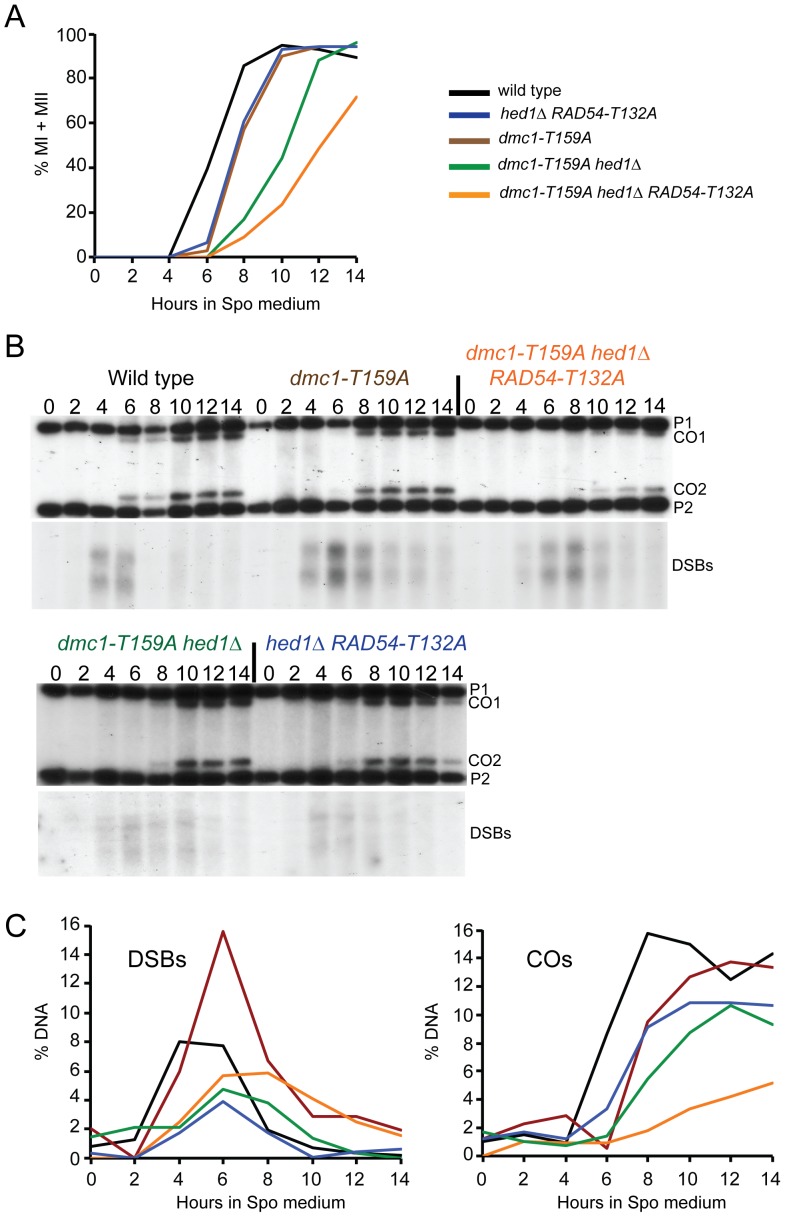
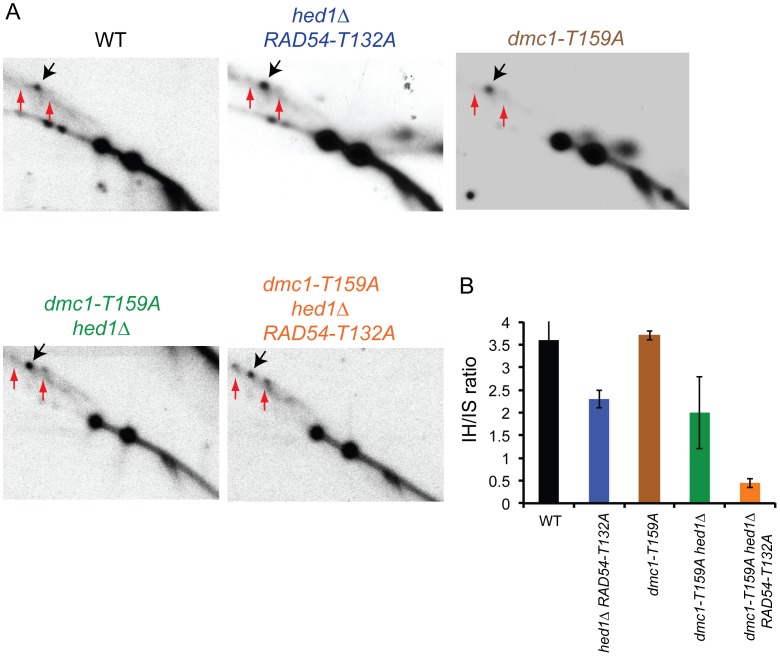
Interhomolog recombination plays a critical role in promoting proper meiotic chromosome segregation but a mechanistic understanding of this process is far from complete. In vegetative cells, Rad51 is a highly conserved recombinase that exhibits a preference for repairing double strand breaks (DSBs) using sister chromatids, in contrast to the conserved, meiosis-specific recombinase, Dmc1, which preferentially repairs programmed DSBs using homologs. Despite the different preferences for repair templates, both Rad51 and Dmc1 are required for interhomolog recombination during meiosis. This paradox has recently been explained by the finding that Rad51 protein, but not its strand exchange activity, promotes Dmc1 function in budding yeast. Rad51 activity is inhibited in dmc1Δ mutants, where the failure to repair meiotic DSBs triggers the meiotic recombination checkpoint, resulting in prophase arrest. The question remains whether inhibition of Rad51 activity is important during wild-type meiosis, or whether inactivation of Rad51 occurs only as a result of the absence of DMC1 or checkpoint activation. This work shows that strains in which mechanisms that down-regulate Rad51 activity are removed exhibit reduced numbers of interhomolog crossovers and noncrossovers. A hypomorphic mutant, dmc1-T159A, makes less stable presynaptic filaments but is still able to mediate strand exchange and interact with accessory factors. Combining dmc1-T159A with up-regulated Rad51 activity reduces interhomolog recombination and spore viability, while increasing intersister joint molecule formation. These results support the idea that down-regulation of Rad51 activity is important during meiosis to prevent Rad51 from competing with Dmc1 for repair of meiotic DSBs.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
The sense and sensibility of strand exchange in recombination homeostasis.PLoS Genet. 2014 Jan;10(1):e1004104. doi: 10.1371/journal.pgen.1004104. Epub 2014 Jan 23. PLoS Genet. 2014. PMID: 24465222 Free PMC article. No abstract available.
References
-
- Petronczki M, Siomos MF, Nasmyth K (2003) Un menage a quatre: the molecular biology of chromosome segregation in meiosis. Cell 112: 423–440. - PubMed
-
- Hassold T, Hunt P (2001) To err (meiotically) is human: the genesis of human aneuploidy. Nature reviews 2: 280–291. - PubMed
-
- Sheridan S, Bishop DK (2006) Red-Hed regulation: recombinase Rad51, though capable of playing the leading role, may be relegated to supporting Dmc1 in budding yeast meiosis. Genes Dev 20: 1685–1691. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
