

The role of flotillins in regulating aβ production, investigated using flotillin 1-/-, flotillin 2-/- double knockout mice
- PMID: 24465508
- PMCID: PMC3897416
- DOI: 10.1371/journal.pone.0085217
The role of flotillins in regulating aβ production, investigated using flotillin 1-/-, flotillin 2-/- double knockout mice
Abstract
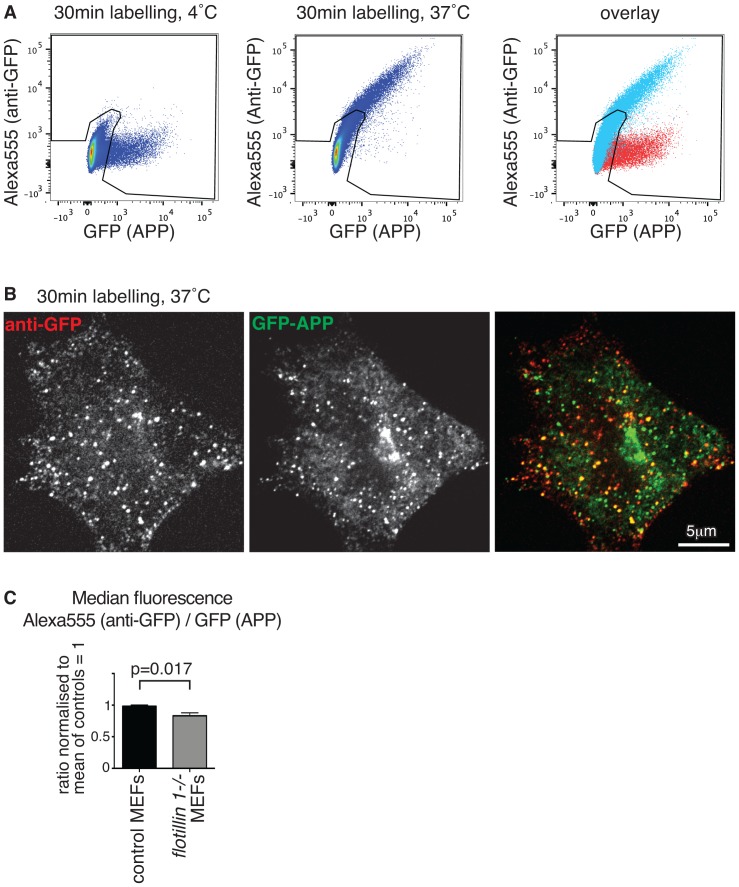
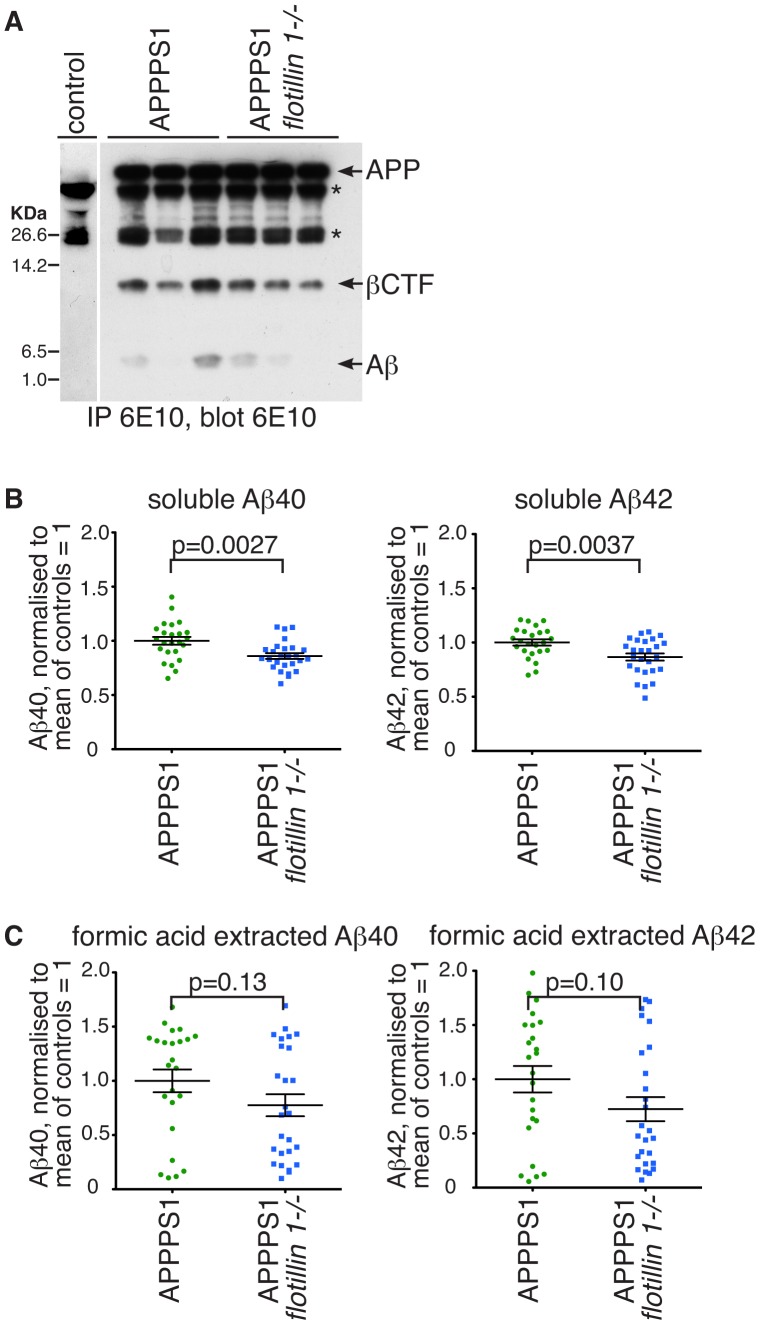
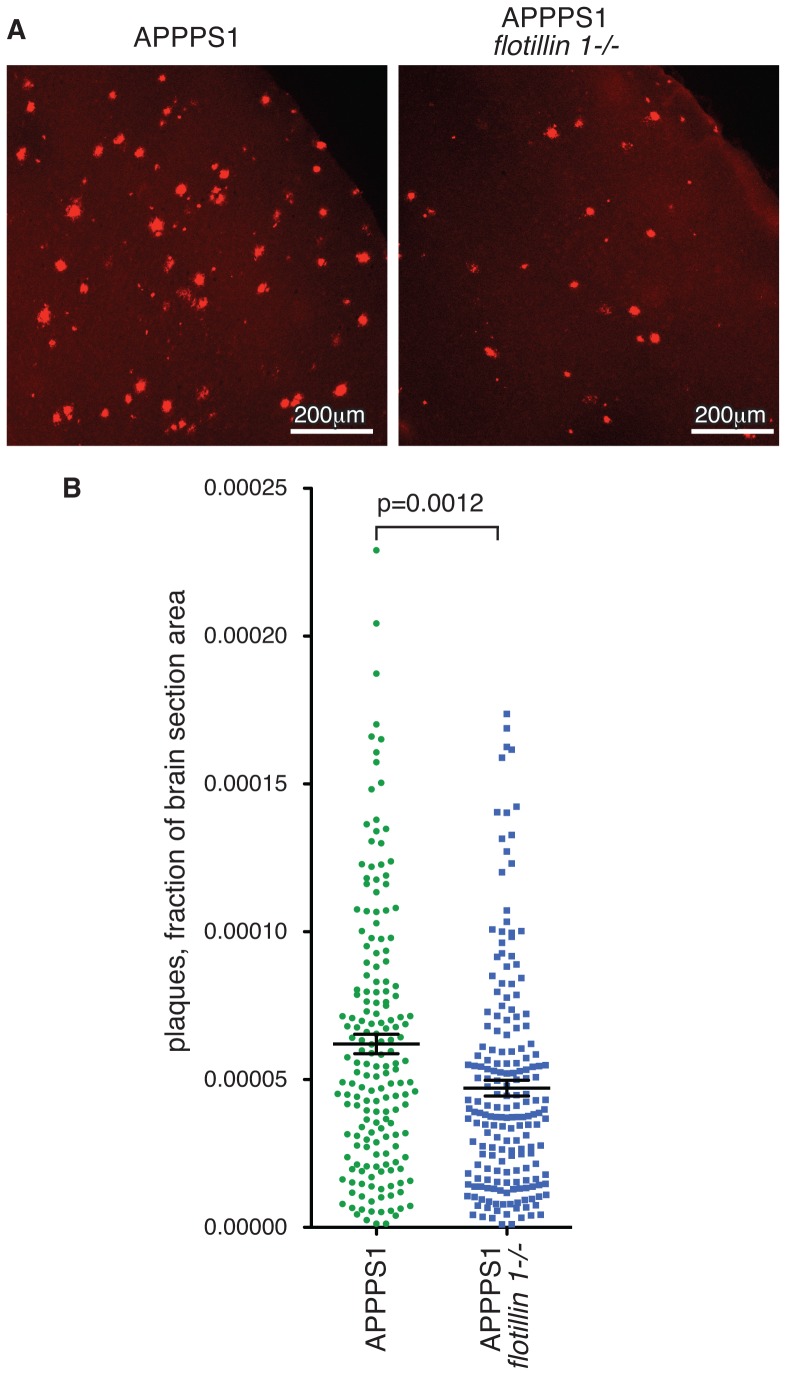
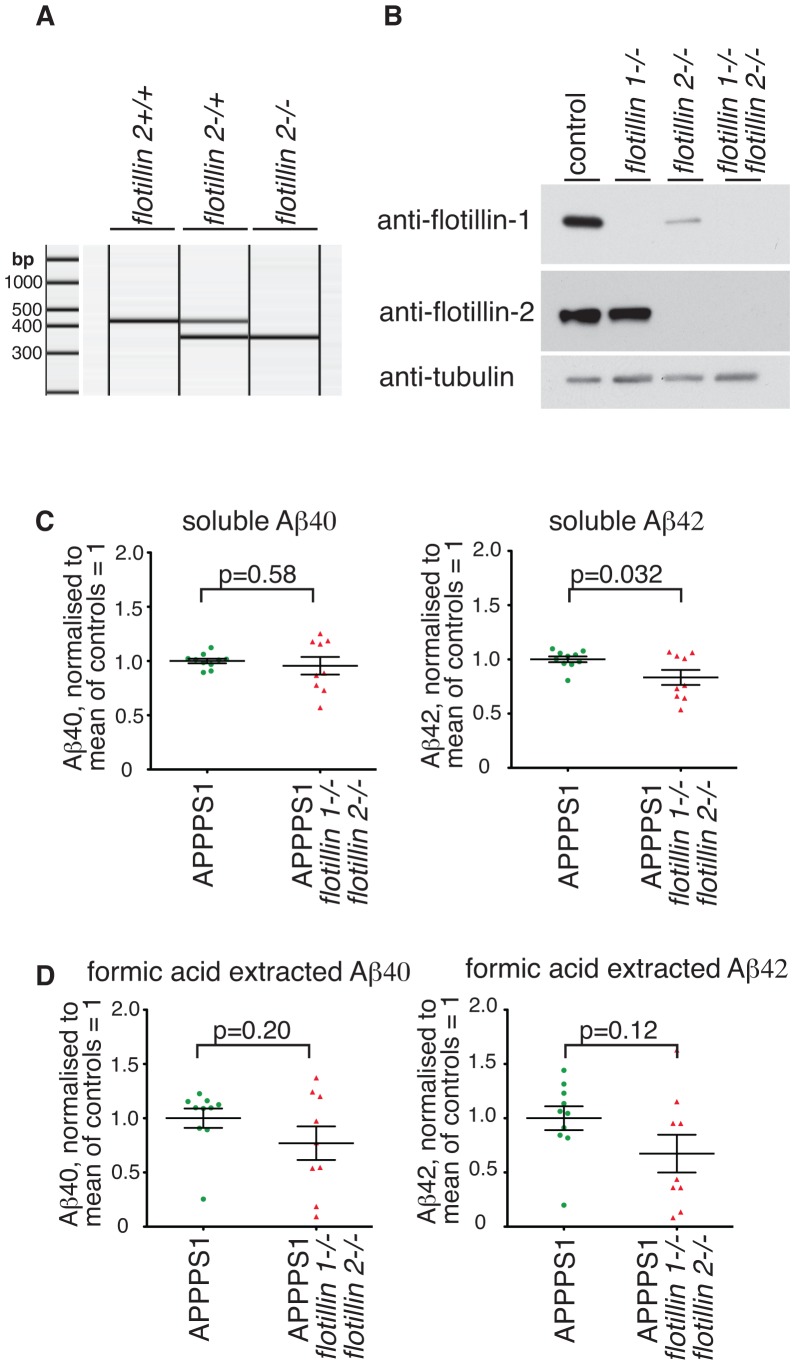
Flotillin 1 and flotillin 2 associate in the plasma membrane to form microdomains that have roles in cell signaling, regulation of cell-cell contacts, membrane-cytoskeletal interactions, and endocytosis. They are thought to be involved in the trafficking and hence processing of the Amyloid Precursor Protein, APP. In this study we set out to obtain in vivo confirmation of a link between flotillins and cleavage of APP to release amyloidogenic Aβ peptide, and to generate tools that would allow us to ask whether flotillins are functionally redundant. We used a mouse model for Aβ-dependent cerebral amyloidosis, APPPS1 mice, combined with deletion of either flotillin 1 singly, or flotillin 1 and flotillin 2 together. There was a small but significant reduction in Aβ levels, and the abundance of congo-red stained plaques, in brains of 12 week old mice lacking flotillin 1. A similar reduction in Aβ levels was observed in the flotillin 1-/-, flotillin 2-/- double knockouts. We did not observe large effects on the clustering or endocytosis of APP in flotillin 1-/- mouse embryonic fibroblasts. We conclude that flotillins are likely to play some role in APP trafficking or processing, but the relevant cellular mechanisms require more investigation. The availability of flotillin 1-/-, flotillin 2-/- mice, which have no overt phenotypes, will facilitate research into flotillin function in vivo.
Conflict of interest statement
Figures

Similar articles
-
Flotillin-dependent clustering of the amyloid precursor protein regulates its endocytosis and amyloidogenic processing in neurons.J Neurosci. 2008 Mar 12;28(11):2874-82. doi: 10.1523/JNEUROSCI.5345-07.2008. J Neurosci. 2008. PMID: 18337418 Free PMC article.
-
Increased activity of mitogen activated protein kinase pathway in flotillin-2 knockout mouse model.Cell Signal. 2014 Feb;26(2):198-207. doi: 10.1016/j.cellsig.2013.11.001. Epub 2013 Nov 9. Cell Signal. 2014. PMID: 24216609
-
Murine versus human apolipoprotein E4: differential facilitation of and co-localization in cerebral amyloid angiopathy and amyloid plaques in APP transgenic mouse models.Acta Neuropathol Commun. 2015 Nov 10;3:70. doi: 10.1186/s40478-015-0250-y. Acta Neuropathol Commun. 2015. PMID: 26556230 Free PMC article.
-
Hereditary and sporadic forms of abeta-cerebrovascular amyloidosis and relevant transgenic mouse models.Int J Mol Sci. 2009 Apr 23;10(4):1872-1895. doi: 10.3390/ijms10041872. Int J Mol Sci. 2009. PMID: 19468344 Free PMC article. Review.
-
Flotillins in membrane trafficking and physiopathology.Biol Cell. 2025 Jan;117(1):e2400134. doi: 10.1111/boc.202400134. Biol Cell. 2025. PMID: 39877933 Free PMC article. Review.
Cited by
-
Neurodegeneration and Neuro-Regeneration-Alzheimer's Disease and Stem Cell Therapy.Int J Mol Sci. 2019 Aug 31;20(17):4272. doi: 10.3390/ijms20174272. Int J Mol Sci. 2019. PMID: 31480448 Free PMC article. Review.
-
Flot2 acts as a novel mediator of podocyte injury in proteinuric kidney disease.Int J Biol Sci. 2023 Jan 1;19(2):502-520. doi: 10.7150/ijbs.78945. eCollection 2023. Int J Biol Sci. 2023. PMID: 36632460 Free PMC article.
-
The Flot2 component of the lipid raft changes localization during neural differentiation of P19C6 cells.BMC Mol Cell Biol. 2019 Aug 27;20(1):38. doi: 10.1186/s12860-019-0225-0. BMC Mol Cell Biol. 2019. PMID: 31455216 Free PMC article.
-
A plasma membrane microdomain compartmentalizes ephrin-generated cAMP signals to prune developing retinal axon arbors.Nat Commun. 2016 Oct 3;7:12896. doi: 10.1038/ncomms12896. Nat Commun. 2016. PMID: 27694812 Free PMC article.
-
Selective endocytosis controls slit diaphragm maintenance and dynamics in Drosophila nephrocytes.Elife. 2022 Jul 25;11:e79037. doi: 10.7554/eLife.79037. Elife. 2022. PMID: 35876643 Free PMC article.
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
