

Crystallographic and biochemical analysis of the mouse poly(ADP-ribose) glycohydrolase
- PMID: 24465839
- PMCID: PMC3897571
- DOI: 10.1371/journal.pone.0086010
Crystallographic and biochemical analysis of the mouse poly(ADP-ribose) glycohydrolase
Abstract
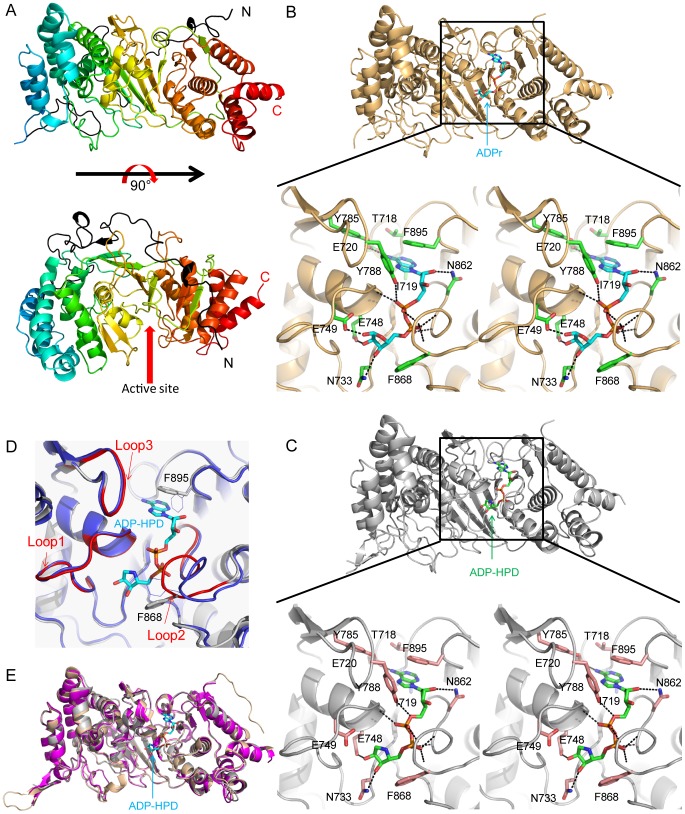
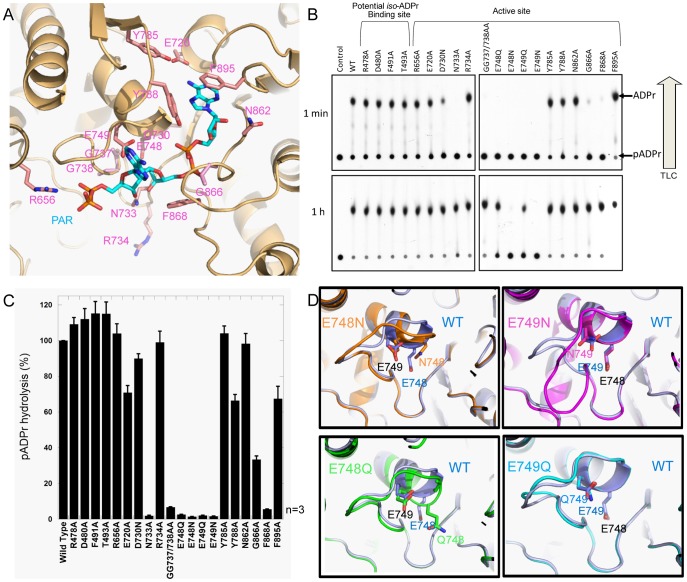
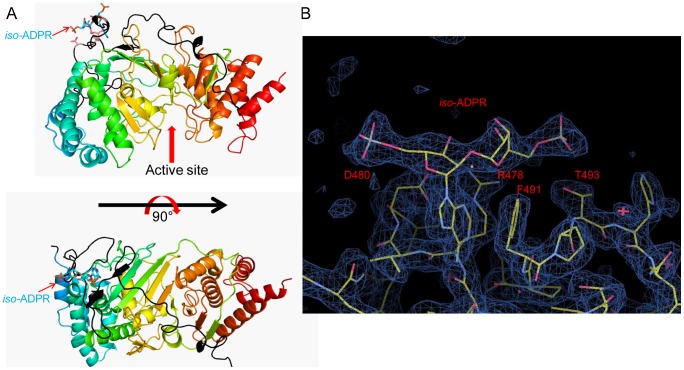
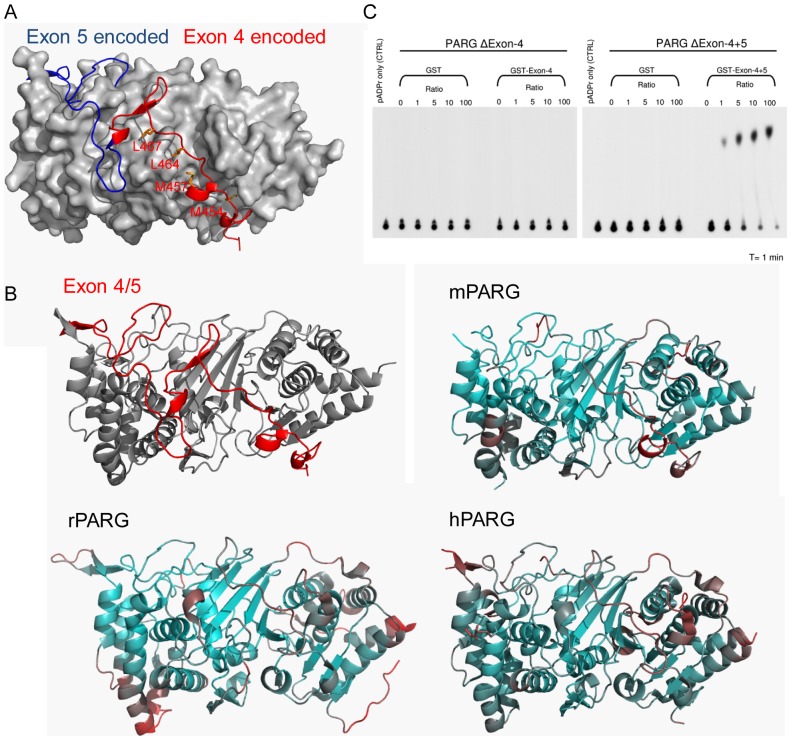
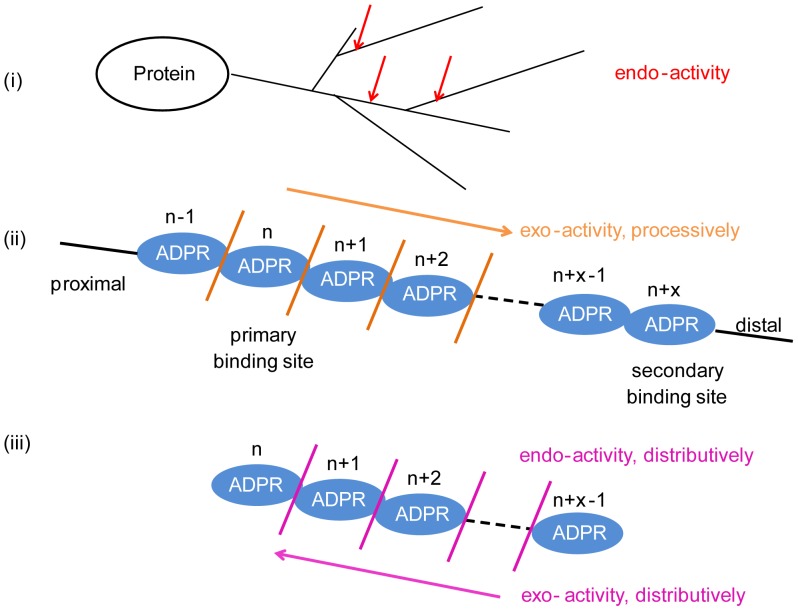
Protein poly(ADP-ribosyl)ation (PARylation) regulates a number of important cellular processes. Poly(ADP-ribose) glycohydrolase (PARG) is the primary enzyme responsible for hydrolyzing the poly(ADP-ribose) (PAR) polymer in vivo. Here we report crystal structures of the mouse PARG (mPARG) catalytic domain, its complexes with ADP-ribose (ADPr) and a PARG inhibitor ADP-HPD, as well as four PARG catalytic residues mutants. With these structures and biochemical analysis of 20 mPARG mutants, we provide a structural basis for understanding how the PAR polymer is recognized and hydrolyzed by mPARG. The structures and activity complementation experiment also suggest how the N-terminal flexible peptide preceding the PARG catalytic domain may regulate the enzymatic activity of PARG. This study contributes to our understanding of PARG catalytic and regulatory mechanisms as well as the rational design of PARG inhibitors.
Conflict of interest statement
Figures
Similar articles
-
The structure and catalytic mechanism of a poly(ADP-ribose) glycohydrolase.Nature. 2011 Sep 4;477(7366):616-20. doi: 10.1038/nature10404. Nature. 2011. PMID: 21892188 Free PMC article.
-
Visualization of poly(ADP-ribose) bound to PARG reveals inherent balance between exo- and endo-glycohydrolase activities.Nat Commun. 2013;4:2164. doi: 10.1038/ncomms3164. Nat Commun. 2013. PMID: 23917065 Free PMC article.
-
Structures of the human poly (ADP-ribose) glycohydrolase catalytic domain confirm catalytic mechanism and explain inhibition by ADP-HPD derivatives.PLoS One. 2012;7(12):e50889. doi: 10.1371/journal.pone.0050889. Epub 2012 Dec 10. PLoS One. 2012. PMID: 23251397 Free PMC article.
-
Poly (ADP-ribose) glycohydrolase (PARG) and its therapeutic potential.Front Biosci (Landmark Ed). 2009 Jan 1;14(5):1619-26. doi: 10.2741/3329. Front Biosci (Landmark Ed). 2009. PMID: 19273151 Review.
-
New Insights into the Roles of NAD+-Poly(ADP-ribose) Metabolism and Poly(ADP-ribose) Glycohydrolase.Curr Protein Pept Sci. 2016;17(7):668-682. doi: 10.2174/1389203717666160419150014. Curr Protein Pept Sci. 2016. PMID: 27817743 Review.
Cited by
-
MacroH2A1 Regulation of Poly(ADP-Ribose) Synthesis and Stability Prevents Necrosis and Promotes DNA Repair.Mol Cell Biol. 2019 Dec 11;40(1):e00230-19. doi: 10.1128/MCB.00230-19. Print 2019 Dec 11. Mol Cell Biol. 2019. PMID: 31636161 Free PMC article.
-
Readers of poly(ADP-ribose): designed to be fit for purpose.Nucleic Acids Res. 2016 Feb 18;44(3):993-1006. doi: 10.1093/nar/gkv1383. Epub 2015 Dec 15. Nucleic Acids Res. 2016. PMID: 26673700 Free PMC article. Review.
-
PARG Mutation Uncovers Critical Structural Determinant for Poly(ADP-Ribose) Hydrolysis and Chromatin Regulation in Embryonic Stem Cells.Cells. 2025 Jul 9;14(14):1049. doi: 10.3390/cells14141049. Cells. 2025. PMID: 40710302 Free PMC article.
-
The SARS-CoV-2 Conserved Macrodomain Is a Mono-ADP-Ribosylhydrolase.J Virol. 2021 Jan 13;95(3):e01969-20. doi: 10.1128/JVI.01969-20. Print 2021 Jan 13. J Virol. 2021. PMID: 33158944 Free PMC article.
-
The rise and fall of poly(ADP-ribose): An enzymatic perspective.DNA Repair (Amst). 2015 Aug;32:10-16. doi: 10.1016/j.dnarep.2015.04.008. Epub 2015 May 1. DNA Repair (Amst). 2015. PMID: 25963443 Free PMC article. Review.
References
-
- De Vos M, Schreiber V, Dantzer F (2012) The diverse roles and clinical relevance of PARPs in DNA damage repair: current state of the art. Biochem Pharmacol 84: 137–146. - PubMed
-
- Satoh MS, Lindahl T (1992) Role of poly(ADP-ribose) formation in DNA repair. Nature 356: 356–358. - PubMed
-
- Hottiger MO, Hassa PO, Luscher B, Schuler H, Koch-Nolte F (2010) Toward a unified nomenclature for mammalian ADP-ribosyltransferases. Trends Biochem Sci 35: 208–219. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
