

Evidence from a mouse model that epithelial cell migration and mesenchymal-epithelial transition contribute to rapid restoration of uterine tissue integrity during menstruation
- PMID: 24466063
- PMCID: PMC3899239
- DOI: 10.1371/journal.pone.0086378
Evidence from a mouse model that epithelial cell migration and mesenchymal-epithelial transition contribute to rapid restoration of uterine tissue integrity during menstruation
Abstract
Background: In women dynamic changes in uterine tissue architecture occur during each menstrual cycle. Menses, characterised by the shedding of the upper functional layer of the endometrium, is the culmination of a cascade of irreversible changes in tissue function including stromal decidualisation, inflammation and production of degradative enzymes. The molecular mechanisms that contribute to the rapid restoration of tissue homeostasis at time of menses are poorly understood.
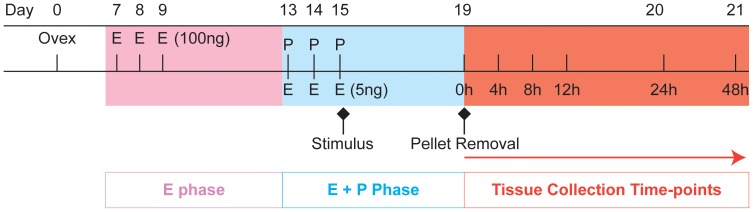
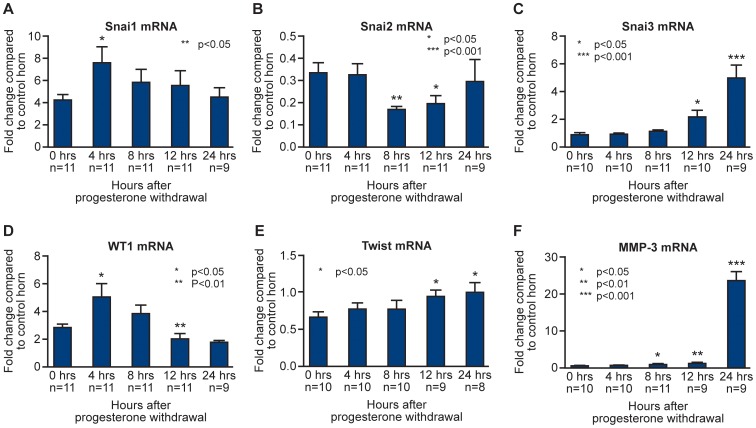
Methodology: A modified mouse model of menses was developed to focus on the events occurring within the uterine lining during endometrial shedding/repair. Decidualisation, vaginal bleeding, tissue architecture and cell proliferation were evaluated at 4, 8, 12, and 24 hours after progesterone (P4) withdrawal; mice received a single injection of bromodeoxyuridine (BrdU) 90 mins before culling. Expression of genes implicated in the regulation of mesenchymal to epithelial transition (MET) was determined using a RT2 PCR profiler array, qRTPCR and bioinformatic analysis.
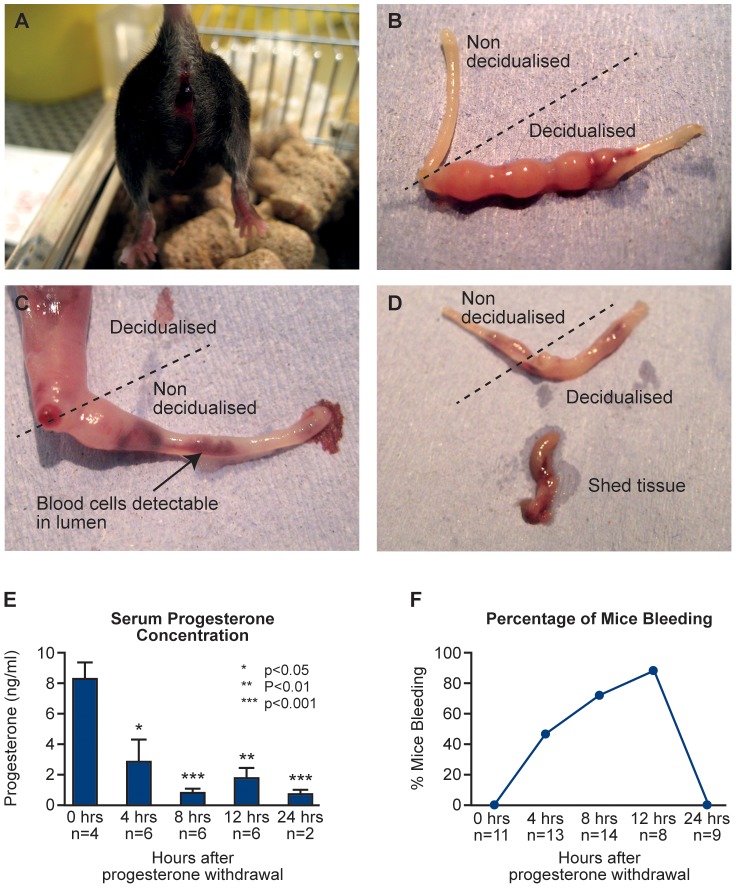
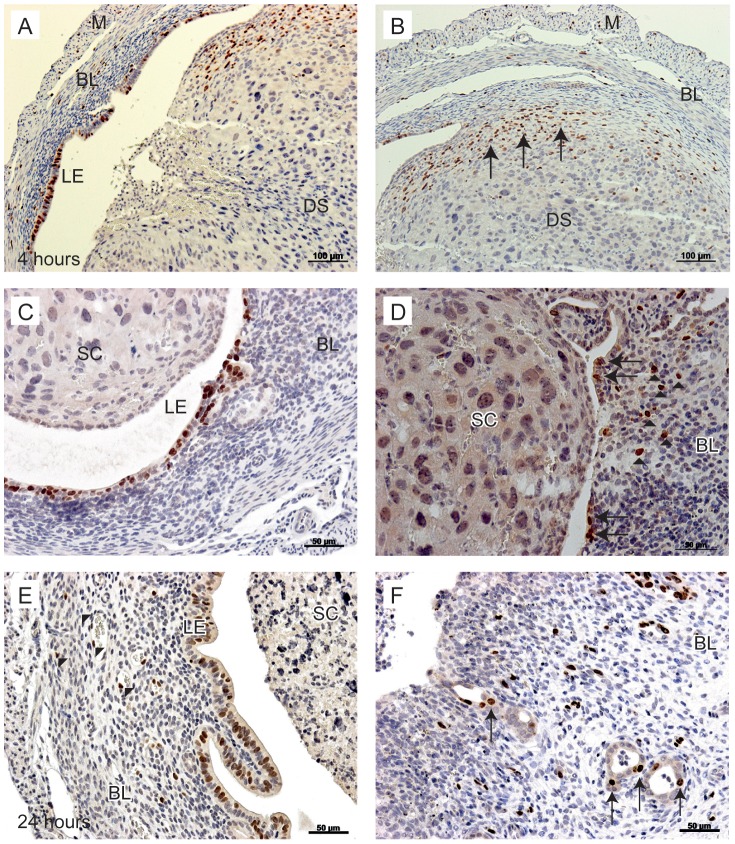
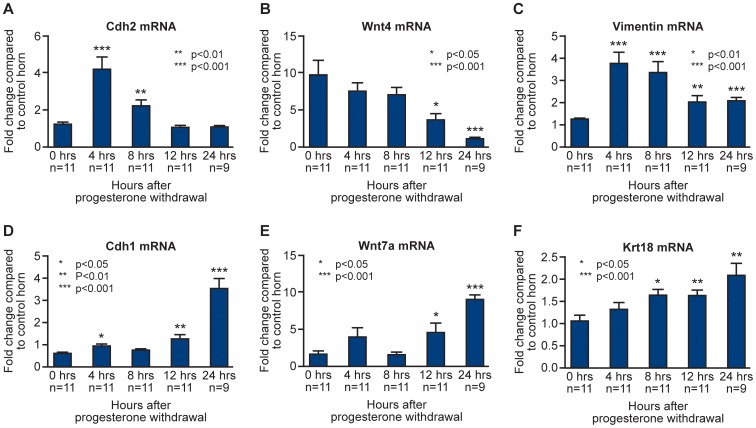
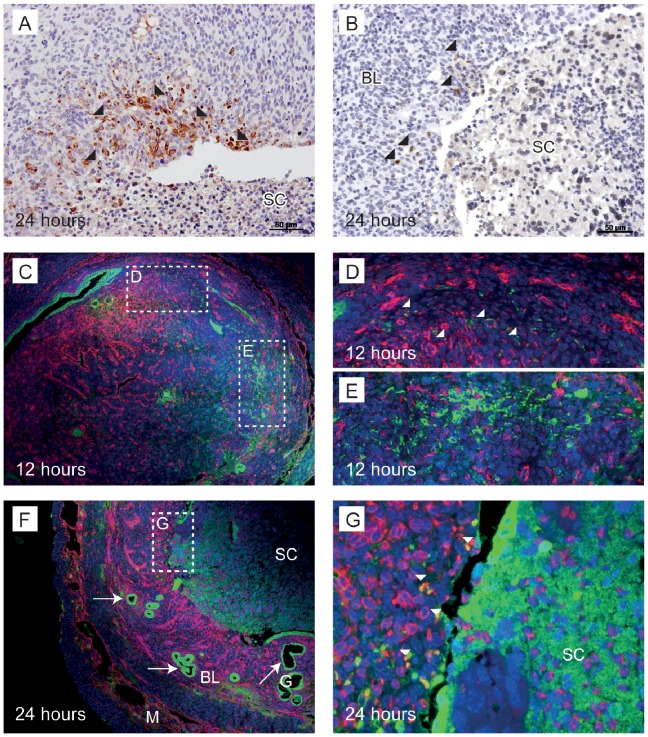
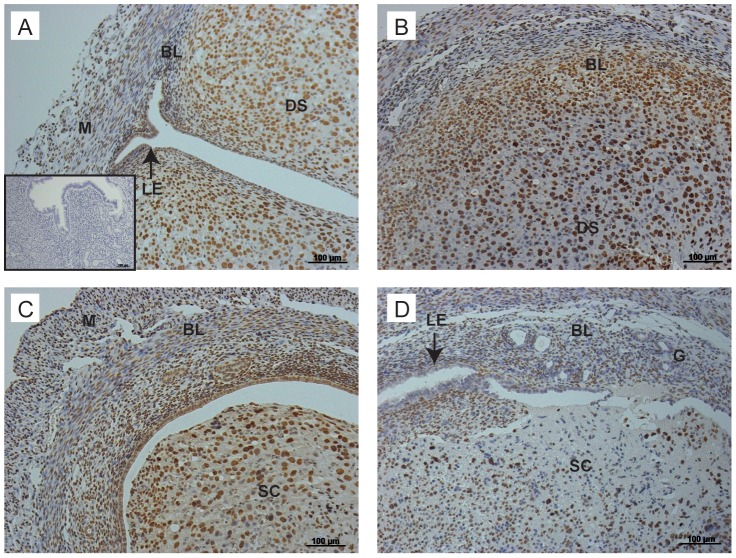
Principal findings: Mice exhibited vaginal bleeding between 4 and 12 hours after P4 withdrawal, concomitant with detachment of the decidualised cell mass from the basal portion of the endometrial lining. Immunostaining for BrdU and pan cytokeratin revealed evidence of epithelial cell proliferation and migration. Cells that appeared to be in transition from a mesenchymal to an epithelial cell identity were identified within the stromal compartment. Analysis of mRNAs encoding genes expressed exclusively in the epithelial or stromal compartments, or implicated in MET, revealed dynamic changes in expression, consistent with a role for reprogramming of mesenchymal cells so that they could contribute to re-epithelialisation.
Conclusions/significance: These studies have provided novel insights into the cellular processes that contribute to re-epithelialisation post-menses implicating both epithelial cell migration and mesenchymal cell differentiation in restoration of an intact epithelial cell layer. These insights may inform development of new therapies to induce rapid healing in the endometrium and other tissues and offer hope to women who suffer from heavy menstrual bleeding.
Conflict of interest statement
Figures

References
-
- Jabbour HN, Kelly RW, Fraser HM, Critchley HO (2006) Endocrine regulation of menstruation. Endocr Rev 27: 17–46. - PubMed
-
- Garry R, Hart R, Karthigasu KA, Burke C (2009) A re-appraisal of the morphological changes within the endometrium during menstruation: a hysteroscopic, histological and scanning electron microscopic study. Hum Reprod 24: 1393–1401. - PubMed
-
- Garry R, Hart R, Karthigasu KA, Burke C (2011) Structural changes in endometrial basal glands during menstruation. BJOG 117: 1175–1185. - PubMed
-
- Ferenczy A (1976) Studies on the cytodynamics of human endometrial regeneration. II. Transmission electron microscopy and histochemistry. Am J Obstet Gynecol 124: 582–595. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
