

Beta-catenin is vital for the integrity of mouse embryonic stem cells
- PMID: 24466203
- PMCID: PMC3897734
- DOI: 10.1371/journal.pone.0086691
Beta-catenin is vital for the integrity of mouse embryonic stem cells
Abstract
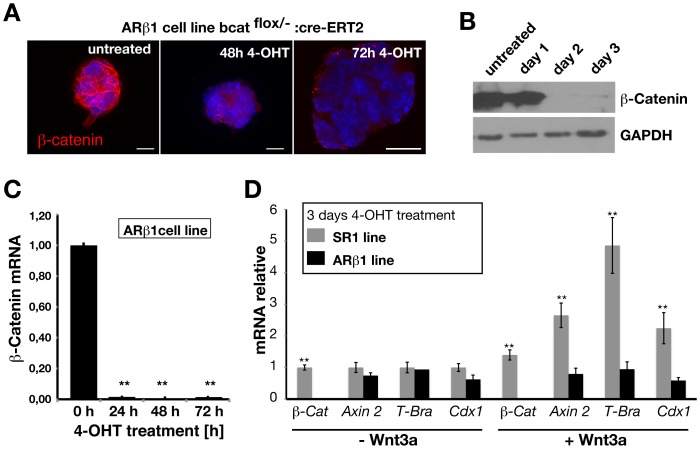
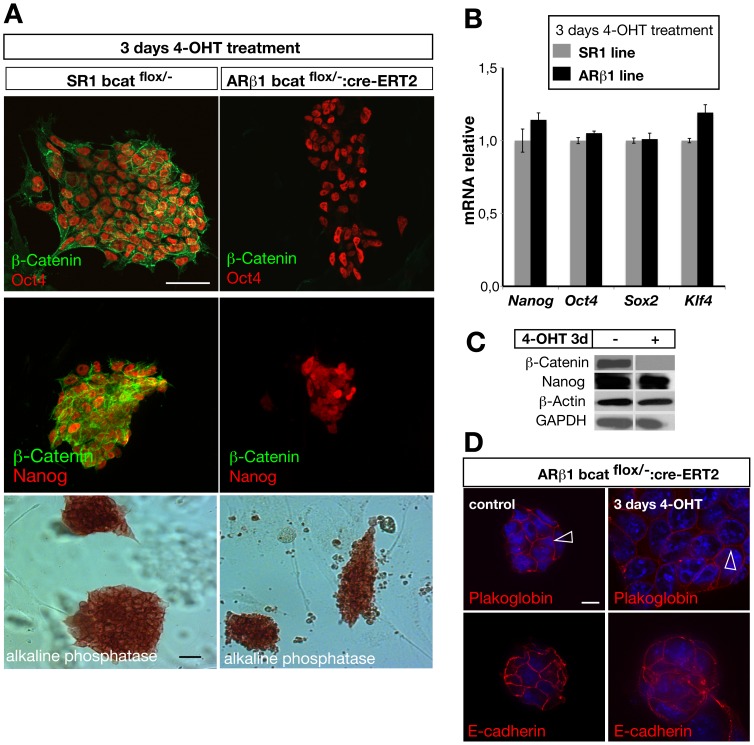
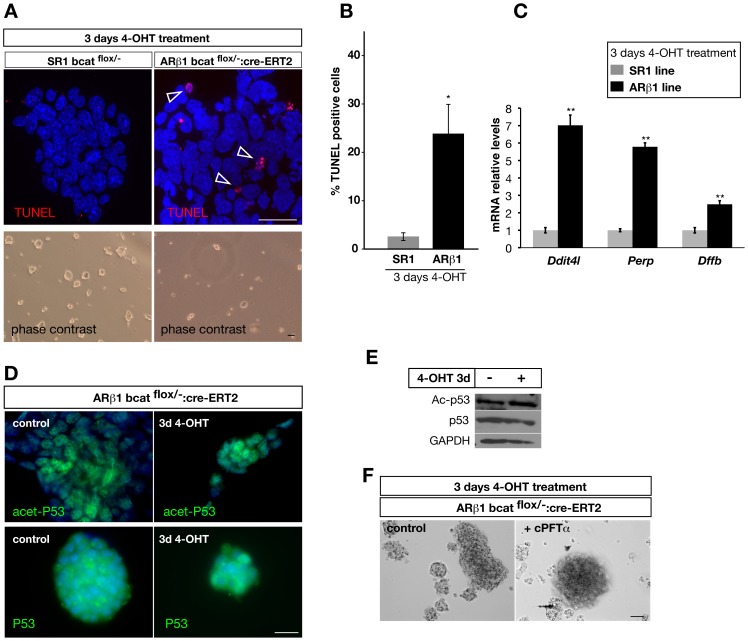
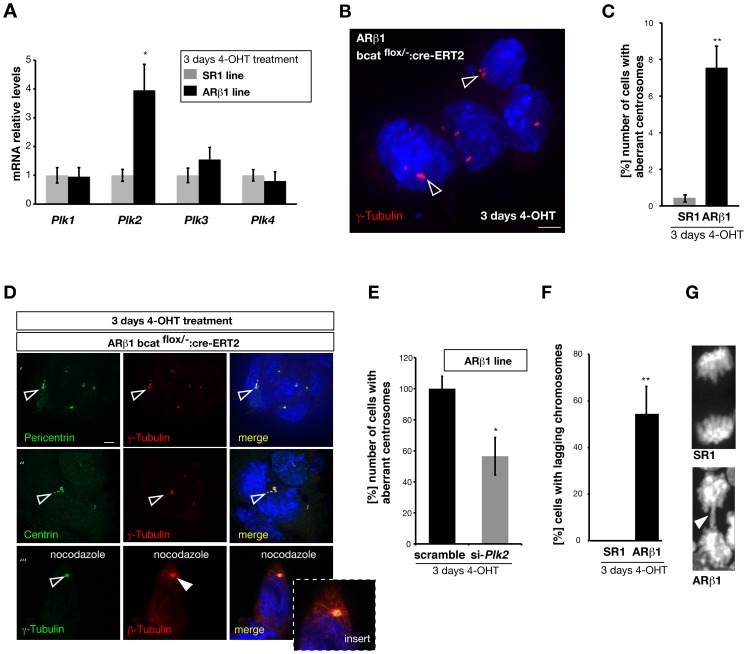
β-Catenin mediated Wnt-signaling is assumed to play a major function in embryonic stem cells in maintaining their stem cell character and the exit from this unique trait. The complexity of β-catenin action and conflicting results on the role of β-catenin in maintaining the pluripotent state have made it difficult to understand its precise cellular and molecular functions. To attempt this issue we have generated new genetically modified mouse embryonic stem cell lines allowing for the deletion of β-catenin in a controlled manner by taking advantage of the Cre-ER-T2 system and analyzed the effects in a narrow time window shortly after ablation. By using this approach, rather then taking long term cultured β-catenin null cell lines we demonstrate that β-catenin is dispensable for the maintenance of pluripotency associated genes. In addition we observed that the removal of β-catenin leads to a strong increase of cell death, the appearance of multiple clustered functional centrosomes most likely due to a mis-regulation of the polo-like-kinase 2 and furthermore, alterations in chromosome segregation. Our study demonstrates the importance of β-catenin in maintaining correct cellular functions and helps to understand its role in embryonic stem cells.
Conflict of interest statement
Figures
References
-
- Evans M (2011) Discovering pluripotency: 30 years of mouse embryonic stem cells. Nat Rev Mol Cell Biol 12: 680–686. - PubMed
-
- Evans MJ, Kaufman MH (1981) Establishment in culture of pluripotential cells from mouse embryos. Nature 292: 154–156. - PubMed
-
- Boiani M, Scholer HR (2005) Regulatory networks in embryo-derived pluripotent stem cells. Nat Rev Mol Cell Biol 6: 872–884. - PubMed
-
- Ng HH, Surani MA (2011) The transcriptional and signalling networks of pluripotency. Nat Cell Biol 13: 490–496. - PubMed
-
- Moon RT, Bowerman B, Boutros M, Perrimon N (2002) The promise and perils of Wnt signaling through beta-catenin. Science 296: 1644–1646. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
