

Epac and the high affinity rolipram binding conformer of PDE4 modulate neurite outgrowth and myelination using an in vitro spinal cord injury model
- PMID: 24467222
- PMCID: PMC3997278
- DOI: 10.1111/bph.12588
Epac and the high affinity rolipram binding conformer of PDE4 modulate neurite outgrowth and myelination using an in vitro spinal cord injury model
Abstract
Background and purpose: cAMP and pharmacological inhibition of PDE4, which degrades it, are promising therapeutic targets for the treatment of spinal cord injury (SCI). Using our previously described in vitro SCI model, we studied the mechanisms by which cAMP modulators promote neurite outgrowth and myelination using enantiomers of the PDE4-specific inhibitor rolipram and other modulators of downstream signalling effectors.
Experimental approach: Rat mixed neural cell myelinating cultures were cut with a scalpel and treated with enantiomers of the PDE4-specific inhibitor rolipram, Epac agonists and PKA antagonists. Neurite outgrowth, density and myelination were assessed by immunocytochemistry and cytokine levels analysed by qPCR.
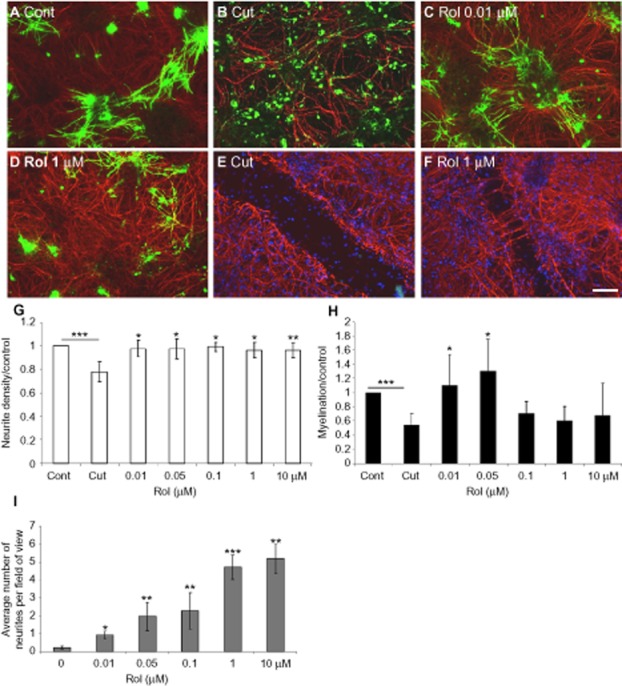
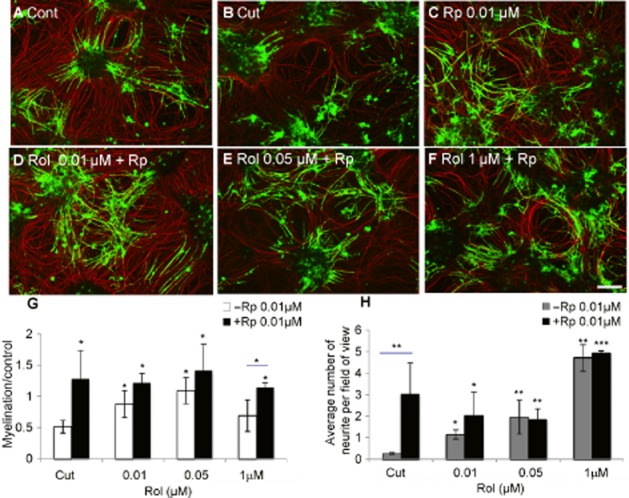
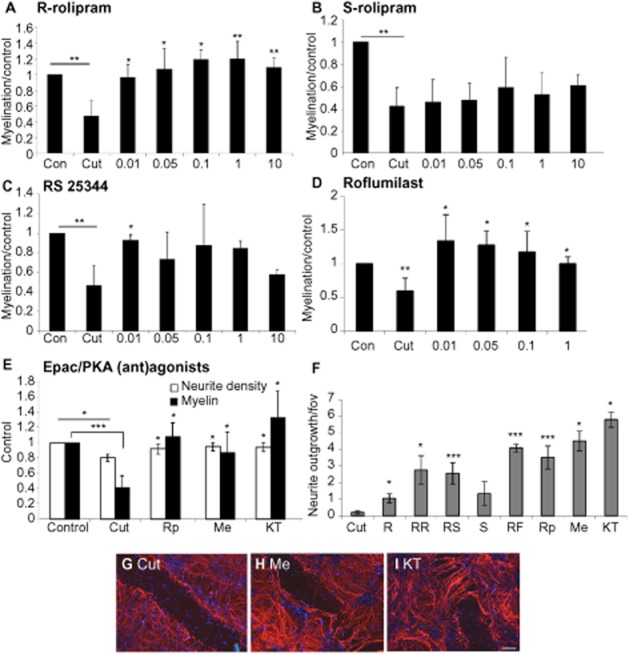
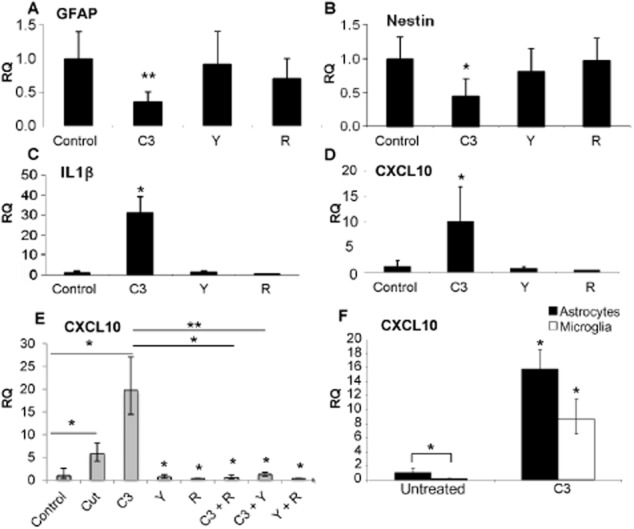
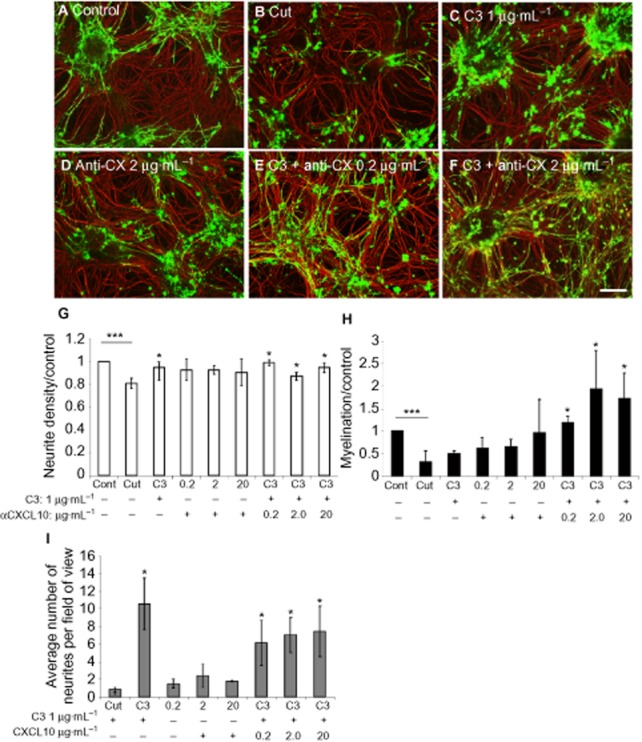
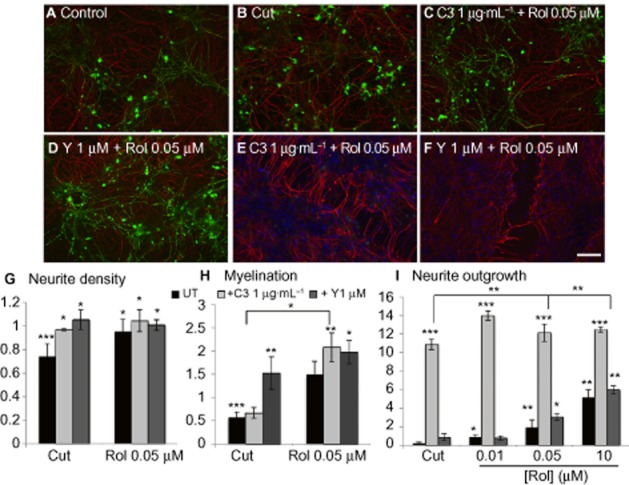
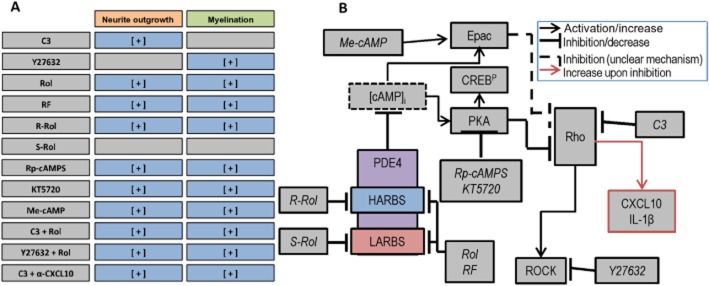
Key results: Inhibition of the high-affinity rolipram-binding state (HARBS), rather than the low-affinity rolipram binding state (LARBS) PDE4 conformer promoted neurite outgrowth and myelination. These effects were mediated through the activation of Epac and not through PKA. Expression of the chemokine CXCL10, known to inhibit myelination, was markedly elevated in astrocytes after Rho inhibition and this was blocked by inhibition of Rho kinase or PDE4.
Conclusions and implications: PDE4 inhibitors targeted at the HARBS conformer or Epac agonists may provide promising novel targets for the treatment of SCI. Our study demonstrates the differential mechanisms of action of these compounds, as well as the benefit of a combined pharmacological approach and highlighting potential promising targets for the treatment of SCI. These findings need to be confirmed in vivo.
Keywords: Epac; PDE4; cAMP; myelination; neurite outgrowth; spinal cord injury.
© 2014 The Authors. British Journal of Pharmacology published by John Wiley & Sons Ltd on behalf of The British Pharmacological Society.
Figures

References
-
- Boomkamp SD, Riehle MO, Wood J, Olson M, Barnett SC. The development of a rat in vitro model of spinal cord injury demonstrating the additive effects of Rho and ROCK inhibitors on neurite outgrowth and myelination. Glia. 2012;3:441–456. - PubMed
-
- Burgin AB, Magnusson OT, Singh J, Witte P, Staker BL, Bjornsson JM, et al. Design of phosphodiesterase 4D (PDE4D) allosteric modulators for enhancing cognition with improved safety. Nat Biotechnol. 2010;28:63–70. - PubMed
-
- Cai D, Shen Y, De Bellard M, Tang S, Filbin MT. Prior exposure to neurotrophins blocks inhibition of axonal regeneration by MAG and myelin via a cAMP-dependent mechanism. Neuron. 1999;22:89–101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
