

INO80-dependent regression of ecdysone-induced transcriptional responses regulates developmental timing in Drosophila
- PMID: 24468295
- PMCID: PMC4005881
- DOI: 10.1016/j.ydbio.2014.01.006
INO80-dependent regression of ecdysone-induced transcriptional responses regulates developmental timing in Drosophila
Abstract
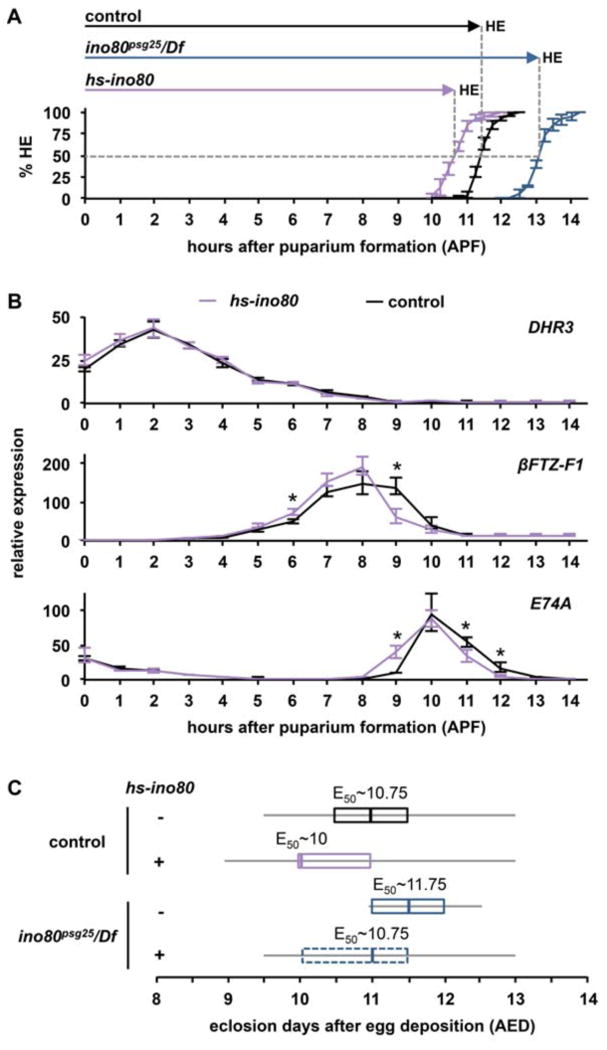
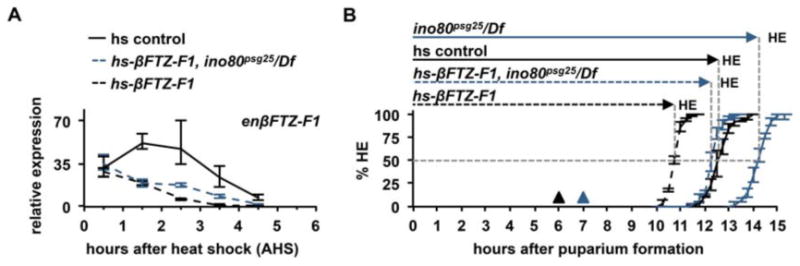
Sequential pulses of the steroid hormone ecdysone regulate the major developmental transitions in Drosophila, and the duration of each developmental stage is determined by the length of time between ecdysone pulses. Ecdysone regulates biological responses by directly initiating target gene transcription. In turn, these transcriptional responses are known to be self-limiting, with mechanisms in place to ensure regression of hormone-dependent transcription. However, the biological significance of these transcriptional repression mechanisms remains unclear. Here we show that the chromatin remodeling protein INO80 facilitates transcriptional repression of ecdysone-regulated genes during prepupal development. In ino80 mutant animals, inefficient repression of transcriptional responses to the late larval ecdysone pulse delays the onset of the subsequent prepupal ecdysone pulse, resulting in a significantly longer prepupal stage. Conversely, increased expression of ino80 is sufficient to shorten the prepupal stage by increasing the rate of transcriptional repression. Furthermore, we demonstrate that enhancing the rate of regression of the mid-prepupal competence factor βFTZ-F1 is sufficient to determine the timing of head eversion and thus the duration of prepupal development. Although ino80 is conserved from yeast to humans, this study represents the first characterization of a bona fide ino80 mutation in any metazoan, raising the possibility that the functions of ino80 in transcriptional repression and developmental timing are evolutionarily conserved.
Keywords: Chromatin remodeler; Developmental timing; Ecdysone; Transcriptional repression.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
DHR3 is required for the prepupal-pupal transition and differentiation of adult structures during Drosophila metamorphosis.Dev Biol. 1999 Aug 1;212(1):204-16. doi: 10.1006/dbio.1999.9343. Dev Biol. 1999. PMID: 10419696
-
Drosophila E93 promotes adult development and suppresses larval responses to ecdysone during metamorphosis.Dev Biol. 2022 Jan;481:104-115. doi: 10.1016/j.ydbio.2021.10.001. Epub 2021 Oct 11. Dev Biol. 2022. PMID: 34648816 Free PMC article.
-
Dynamic feedback circuits function as a switch for shaping a maturation-inducing steroid pulse in Drosophila.Development. 2013 Dec;140(23):4730-9. doi: 10.1242/dev.099739. Epub 2013 Oct 30. Development. 2013. PMID: 24173800 Free PMC article.
-
Ecdysone receptors and their biological actions.Vitam Horm. 2000;60:1-73. doi: 10.1016/s0083-6729(00)60016-x. Vitam Horm. 2000. PMID: 11037621 Review. No abstract available.
-
What goes up must come down: transcription factors have their say in making ecdysone pulses.Curr Top Dev Biol. 2013;103:35-71. doi: 10.1016/B978-0-12-385979-2.00002-2. Curr Top Dev Biol. 2013. PMID: 23347515 Review.
Cited by
-
Distinct requirements for Pho, Sfmbt, and Ino80 for cell survival in Drosophila.Genetics. 2021 Aug 26;219(1):iyab096. doi: 10.1093/genetics/iyab096. Genetics. 2021. PMID: 34849913 Free PMC article.
-
Evolutionarily Conserved Roles for Blood-Brain Barrier Xenobiotic Transporters in Endogenous Steroid Partitioning and Behavior.Cell Rep. 2017 Oct 31;21(5):1304-1316. doi: 10.1016/j.celrep.2017.10.026. Cell Rep. 2017. PMID: 29091768 Free PMC article.
-
INO80 represses osmostress induced gene expression by resetting promoter proximal nucleosomes.Nucleic Acids Res. 2017 Apr 20;45(7):3752-3766. doi: 10.1093/nar/gkw1292. Nucleic Acids Res. 2017. PMID: 28025392 Free PMC article.
-
Interaction of the Chromatin Remodeling Protein hINO80 with DNA.PLoS One. 2016 Jul 18;11(7):e0159370. doi: 10.1371/journal.pone.0159370. eCollection 2016. PLoS One. 2016. PMID: 27428271 Free PMC article.
-
The INO80 remodeller in transcription, replication and repair.Philos Trans R Soc Lond B Biol Sci. 2017 Oct 5;372(1731):20160290. doi: 10.1098/rstb.2016.0290. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 28847827 Free PMC article. Review.
References
-
- Ambros V. Control of developmental timing in Caenorhabditis elegans. Current Opinion in Genetics & Development. 2000;10:428–433. - PubMed
-
- Andres AJ, Thummel CS. Methods for quantitative analysis of transcription in larvae and prepupae. Methods Cell Biol. 1994;44:565–573. - PubMed
-
- Ashburner M, Chihara C, Meltzer P, Richards G. Temporal control of puffing activity in polytene chromosomes. Cold Spring Harb. Symp Quant Biol. 1974;38:655–662. - PubMed
-
- Bainbridge SP, Bownes M. Staging the metamorphosis of Drosophila melanogaster. Journal of embryology and experimental morphology. 1981;66:57–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
