

C2cd3 is critical for centriolar distal appendage assembly and ciliary vesicle docking in mammals
- PMID: 24469809
- PMCID: PMC3926046
- DOI: 10.1073/pnas.1318737111
C2cd3 is critical for centriolar distal appendage assembly and ciliary vesicle docking in mammals
Abstract
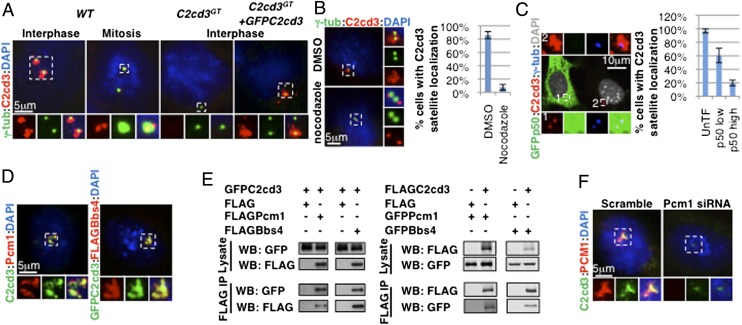
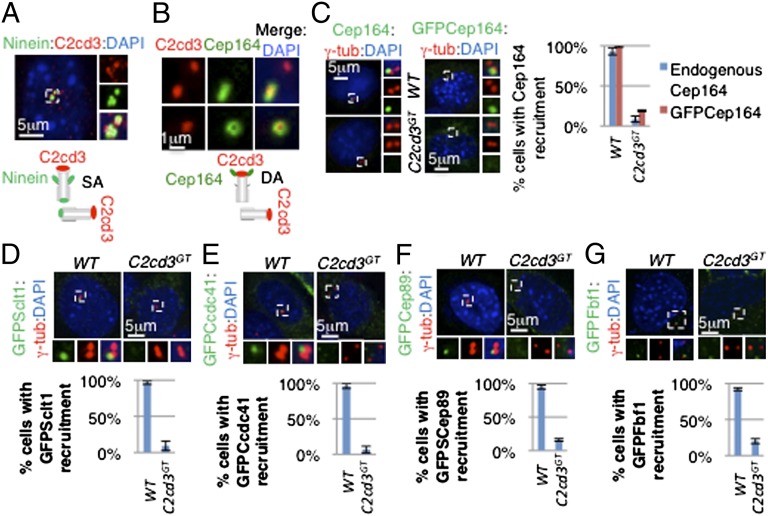
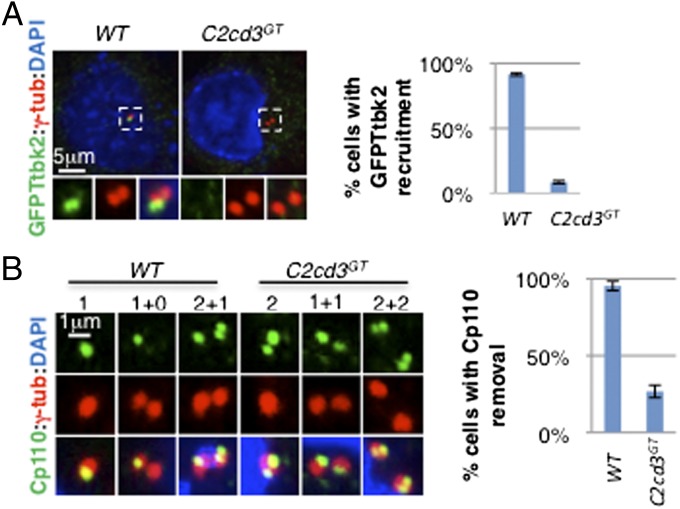
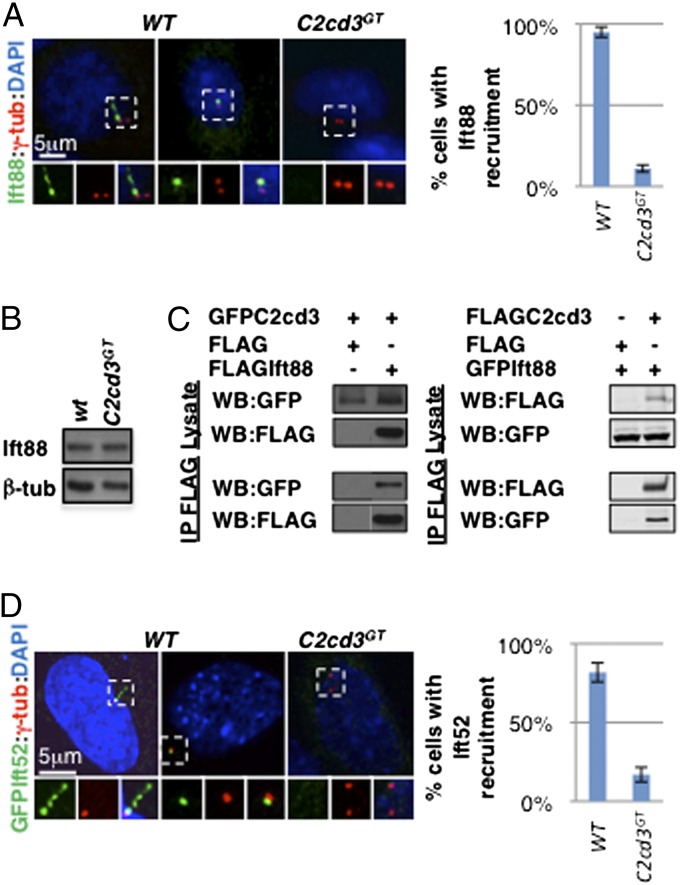
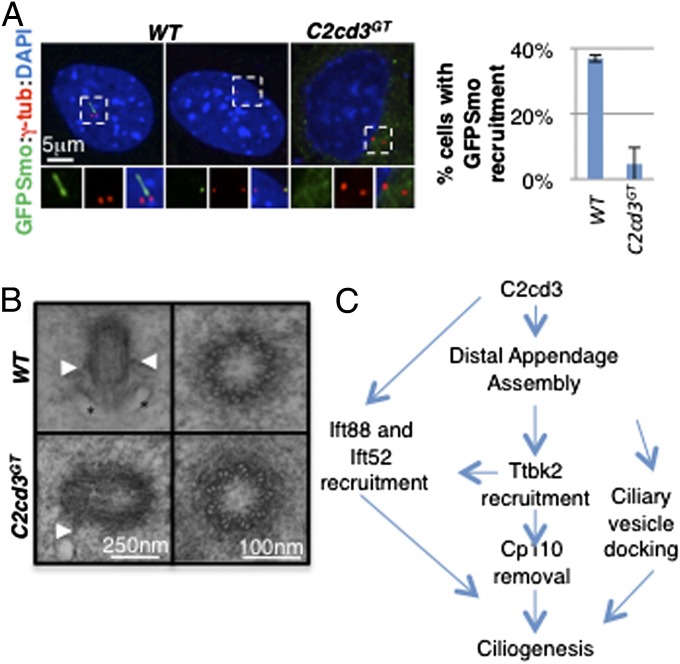
The primary cilium plays critical roles in vertebrate development and physiology, but the mechanisms underlying its biogenesis remain poorly understood. We investigated the molecular function of C2 calcium-dependent domain containing 3 (C2cd3), an essential regulator of primary cilium biogenesis. We show that C2cd3 is localized to the centriolar satellites in a microtubule- and Pcm1-dependent manner; however, C2cd3 is dispensable for centriolar satellite integrity. C2cd3 is also localized to the distal ends of both mother and daughter centrioles and is required for the recruitment of five centriolar distal appendage proteins: Sclt1, Ccdc41, Cep89, Fbf1, and Cep164. Furthermore, loss of C2cd3 results in failure in the recruitment of Ttbk2 to the ciliary basal body as well as the removal of Cp110 from the ciliary basal body, two critical steps in initiating ciliogenesis. C2cd3 is also required for recruiting the intraflagellar transport proteins Ift88 and Ift52 to the mother centriole. Consistent with a role in distal appendage assembly, C2cd3 is essential for ciliary vesicle docking to the mother centriole. Our results suggest that C2cd3 regulates cilium biogenesis by promoting the assembly of centriolar distal appendages critical for docking ciliary vesicles and recruiting other essential ciliogenic proteins.
Keywords: Bbs4; Ofd1; centrosome; ciliopathy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
WDR8 is a centriolar satellite and centriole-associated protein that promotes ciliary vesicle docking during ciliogenesis.J Cell Sci. 2016 Feb 1;129(3):621-36. doi: 10.1242/jcs.179713. Epub 2015 Dec 16. J Cell Sci. 2016. PMID: 26675238
-
CCDC41 is required for ciliary vesicle docking to the mother centriole.Proc Natl Acad Sci U S A. 2013 Apr 9;110(15):5987-92. doi: 10.1073/pnas.1220927110. Epub 2013 Mar 25. Proc Natl Acad Sci U S A. 2013. PMID: 23530209 Free PMC article.
-
A hierarchical pathway for assembly of the distal appendages that organize primary cilia.Elife. 2025 Jan 30;14:e85999. doi: 10.7554/eLife.85999. Elife. 2025. PMID: 39882846 Free PMC article.
-
Accessorizing the centrosome: new insights into centriolar appendages and satellites.Curr Opin Struct Biol. 2021 Feb;66:148-155. doi: 10.1016/j.sbi.2020.10.021. Epub 2020 Dec 3. Curr Opin Struct Biol. 2021. PMID: 33279729 Review.
-
Emerging insights into CP110 removal during early steps of ciliogenesis.J Cell Sci. 2024 Feb 15;137(4):jcs261579. doi: 10.1242/jcs.261579. Epub 2024 Feb 28. J Cell Sci. 2024. PMID: 38415788 Free PMC article. Review.
Cited by
-
A primer on the mouse basal body.Cilia. 2016 Apr 25;5:17. doi: 10.1186/s13630-016-0038-0. eCollection 2016. Cilia. 2016. PMID: 27114821 Free PMC article. Review.
-
The human phosphatase CDC14A modulates primary cilium length by regulating centrosomal actin nucleation.EMBO Rep. 2019 Jan;20(1):e46544. doi: 10.15252/embr.201846544. Epub 2018 Nov 22. EMBO Rep. 2019. PMID: 30467237 Free PMC article.
-
Insights into photoreceptor ciliogenesis revealed by animal models.Prog Retin Eye Res. 2019 Jul;71:26-56. doi: 10.1016/j.preteyeres.2018.12.004. Epub 2018 Dec 25. Prog Retin Eye Res. 2019. PMID: 30590118 Free PMC article. Review.
-
CEP164 Deficiency Causes Hyperproliferation of Pancreatic Cancer Cells.Front Cell Dev Biol. 2020 Nov 5;8:587691. doi: 10.3389/fcell.2020.587691. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33251215 Free PMC article.
-
The small GTPase RSG1 controls a final step in primary cilia initiation.J Cell Biol. 2018 Jan 2;217(1):413-427. doi: 10.1083/jcb.201604048. Epub 2017 Oct 16. J Cell Biol. 2018. PMID: 29038301 Free PMC article.
References
-
- Gerdes JM, Katsanis N. Ciliary function and Wnt signal modulation. Curr Top Dev Biol. 2008;85:175–195. - PubMed
-
- Schneider L, et al. PDGFRalphaalpha signaling is regulated through the primary cilium in fibroblasts. Curr Biol. 2005;15(20):1861–1866. - PubMed
-
- Inglis PN, Boroevich KA, Leroux MR. Piecing together a ciliome. Trends Genet. 2006;22(9):491–500. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
