

Inflammasome activation has an important role in the development of spontaneous colitis
- PMID: 24472848
- PMCID: PMC4115056
- DOI: 10.1038/mi.2014.1
Inflammasome activation has an important role in the development of spontaneous colitis
Abstract
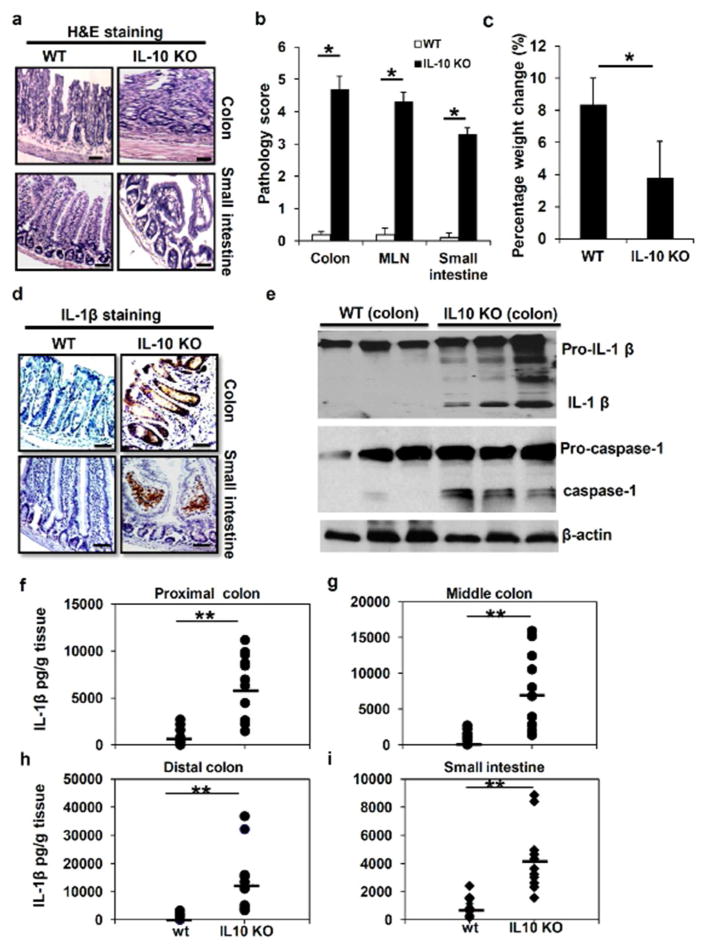
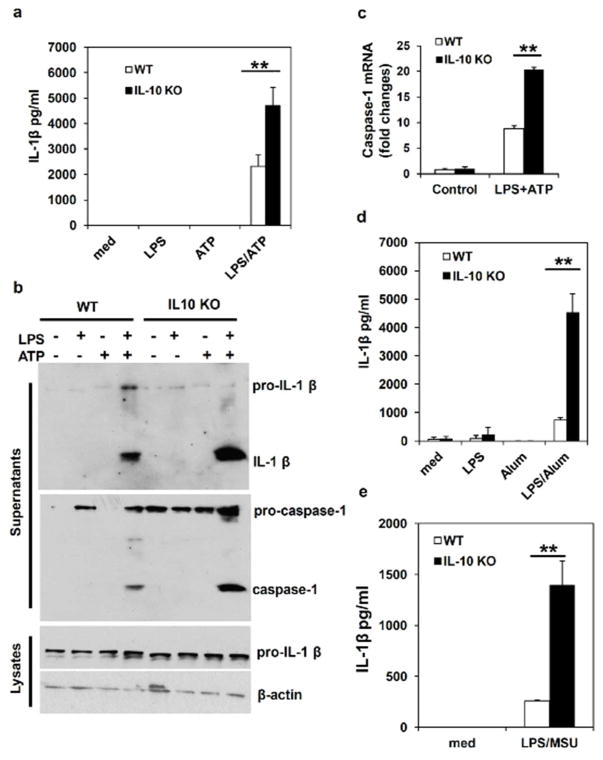
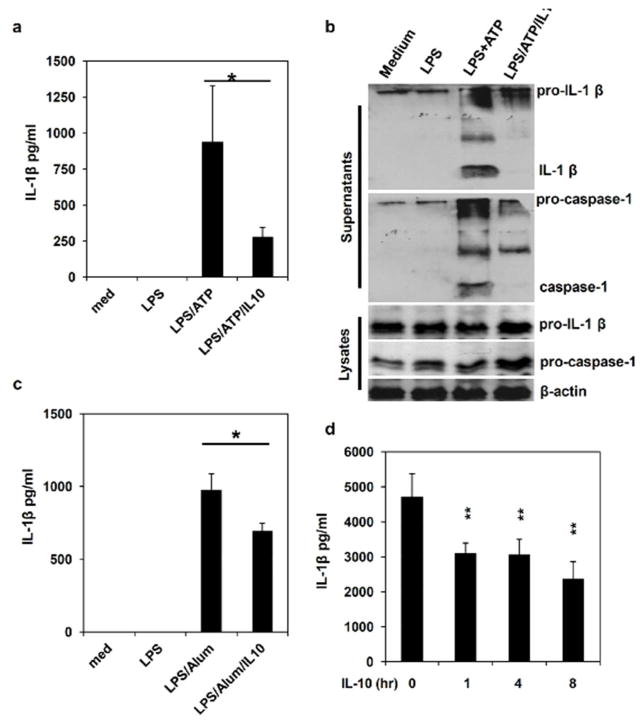
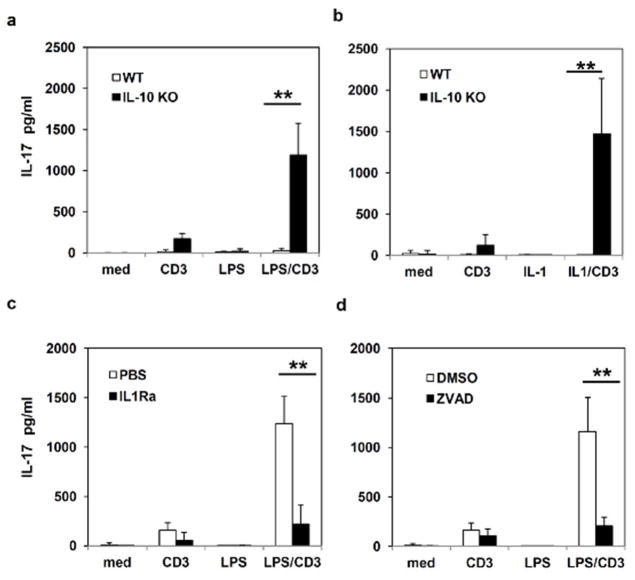
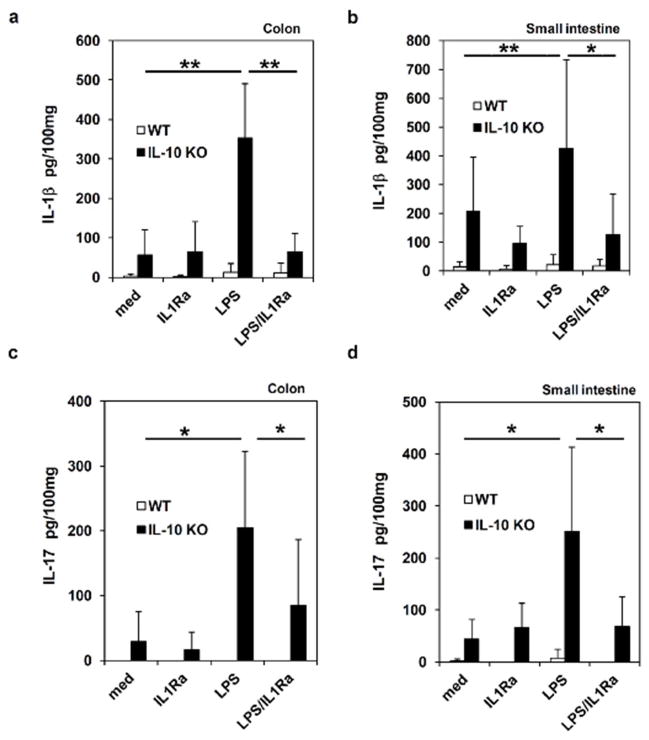
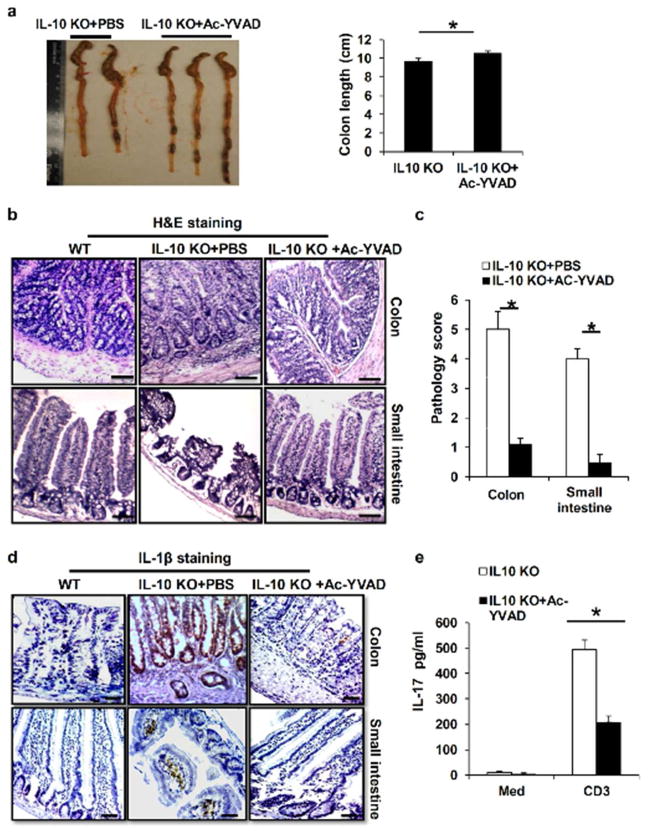
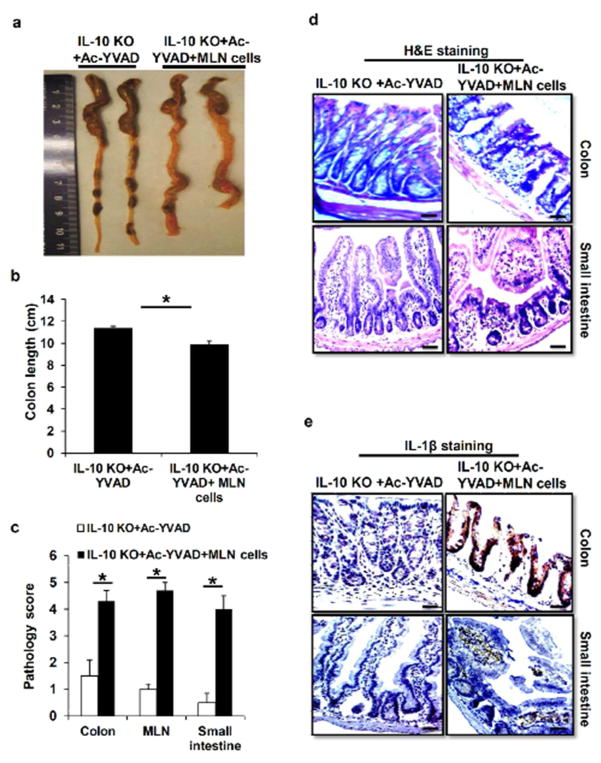
Inflammatory bowel disease (IBD) is characterized for dysregulated intestinal inflammation. Conflicting reports have shown that activation of inflammasome could promote or decrease intestinal inflammation in an acute colitis model, whereas the involvement of inflammasome activation in chronic colitis is poorly understood. In this study, we investigated the role of inflammasome activation in the development of chronic intestinal inflammation by utilizing interleukin-10 (IL-10) knockout (KO) mouse as an animal model, which develops chronic colitis resembling human IBD. We demonstrate the causative link between inflammasome activation and the development of chronic intestinal inflammation. Our results show that mature IL-1β protein levels were significantly increased in all colon sections from IL-10-deficient mice compared with that of wild-type mice. We found that inhibition of inflammasome activities with IL-1 receptor antagonist or caspase-1 inhibitors suppressed IL-1β and IL-17 production from inflamed colon explants. Furthermore, blocking inflammasome activation with caspase-1 inhibitor in vivo significantly ameliorated the spontaneous colitis in IL-10 KO mice. Taken together, these observations demonstrate that inflammasome activation promotes the development of chronic intestinal inflammation.
Conflict of interest statement
Figures
References
-
- Abraham C, Cho J. Interleukin-23/Th17 pathways and inflammatory bowel disease. Inflamm Bowel Dis. 2009;15:1090–1100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
