

O-GlcNAcylation regulates EZH2 protein stability and function
- PMID: 24474760
- PMCID: PMC3910655
- DOI: 10.1073/pnas.1323226111
O-GlcNAcylation regulates EZH2 protein stability and function
Abstract
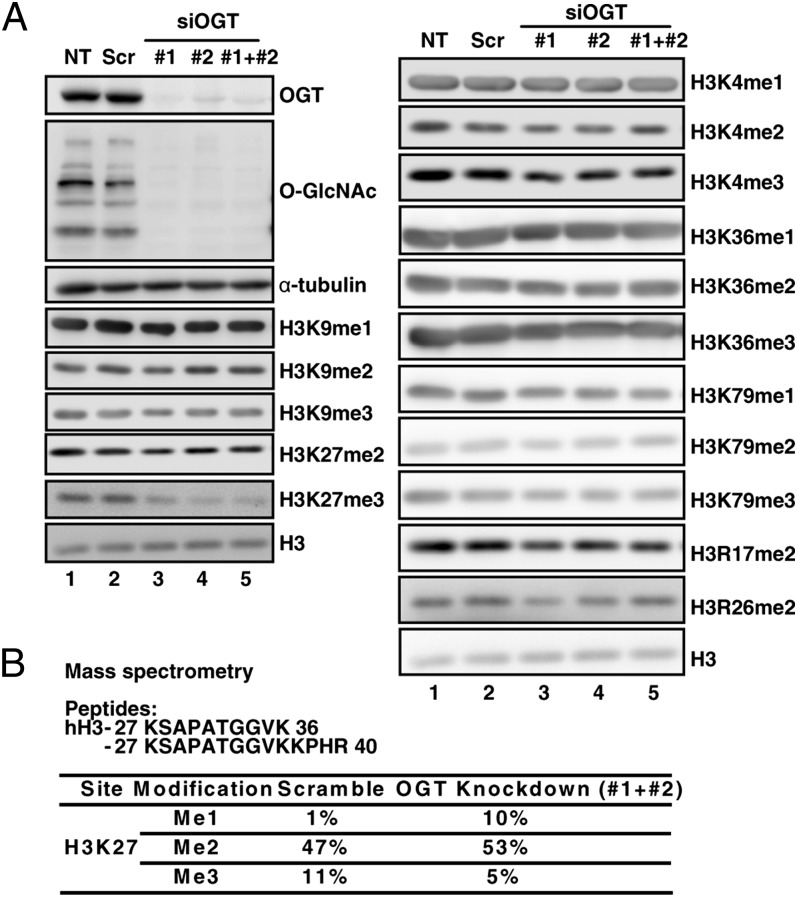
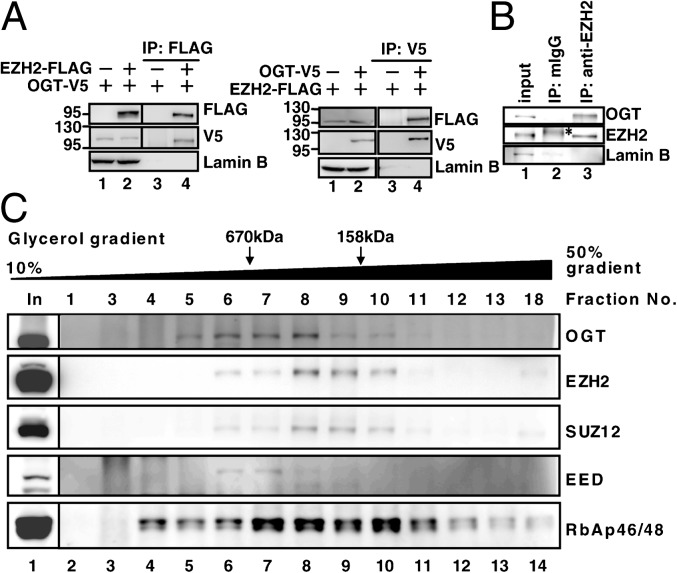
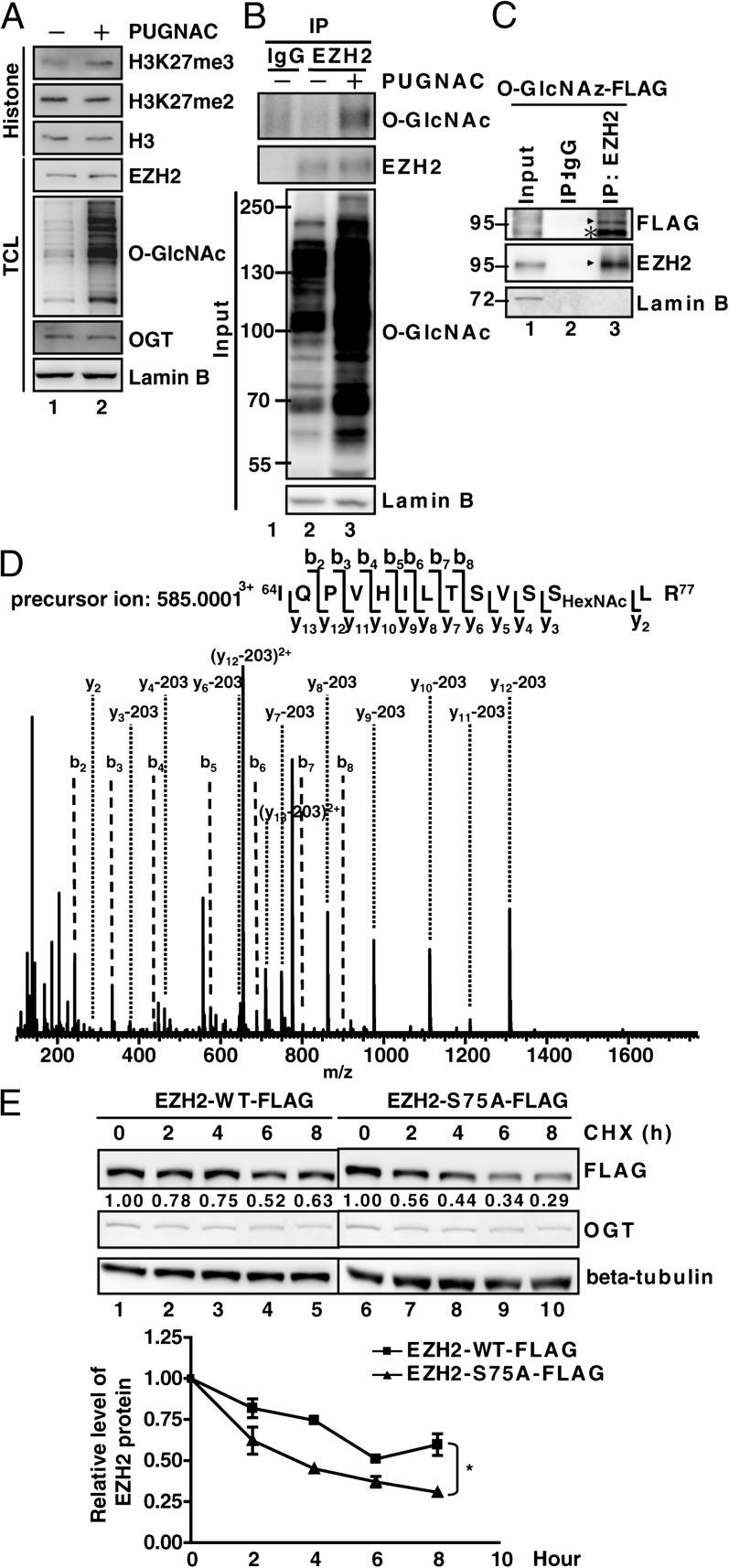
O-linked N-acetylglucosamine (GlcNAc) transferase (OGT) is the only known enzyme that catalyzes the O-GlcNAcylation of proteins at the Ser or Thr side chain hydroxyl group. OGT participates in transcriptional and epigenetic regulation, and dysregulation of OGT has been implicated in diseases such as cancer. However, the underlying mechanism is largely unknown. Here we show that OGT is required for the trimethylation of histone 3 at K27 to form the product H3K27me3, a process catalyzed by the histone methyltransferase enhancer of zeste homolog 2 (EZH2) in the polycomb repressive complex 2 (PRC2). H3K27me3 is one of the most important histone modifications to mark the transcriptionally silenced chromatin. We found that the level of H3K27me3, but not other H3 methylation products, was greatly reduced upon OGT depletion. OGT knockdown specifically down-regulated the protein stability of EZH2, without altering the levels of H3K27 demethylases UTX and JMJD3, and disrupted the integrity of the PRC2 complex. Furthermore, the interaction of OGT and EZH2/PRC2 was detected by coimmunoprecipitation and cosedimentation experiments. Importantly, we identified that serine 75 is the site for EZH2 O-GlcNAcylation, and the EZH2 mutant S75A exhibited reduction in stability. Finally, microarray and ChIP analysis have characterized a specific subset of potential tumor suppressor genes subject to repression via the OGT-EZH2 axis. Together these results indicate that OGT-mediated O-GlcNAcylation at S75 stabilizes EZH2 and hence facilitates the formation of H3K27me3. The study not only uncovers a functional posttranslational modification of EZH2 but also reveals a unique epigenetic role of OGT in regulating histone methylation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Torres CR, Hart GW. Topography and polypeptide distribution of terminal N-acetylglucosamine residues on the surfaces of intact lymphocytes. Evidence for O-linked GlcNAc. J Biol Chem. 1984;259(5):3308–3317. - PubMed
-
- Holt GD, Hart GW. The subcellular distribution of terminal N-acetylglucosamine moieties. Localization of a novel protein-saccharide linkage, O-linked GlcNAc. J Biol Chem. 1986;261(17):8049–8057. - PubMed
-
- Haltiwanger RS, Holt GD, Hart GW. Enzymatic addition of O-GlcNAc to nuclear and cytoplasmic proteins. Identification of a uridine diphospho-N-acetylglucosamine:peptide beta-N-acetylglucosaminyltransferase. J Biol Chem. 1990;265(5):2563–2568. - PubMed
-
- Kreppel LK, Blomberg MA, Hart GW. Dynamic glycosylation of nuclear and cytosolic proteins. Cloning and characterization of a unique O-GlcNAc transferase with multiple tetratricopeptide repeats. J Biol Chem. 1997;272(14):9308–9315. - PubMed
-
- Dong DL, Hart GW. Purification and characterization of an O-GlcNAc selective N-acetyl-beta-D-glucosaminidase from rat spleen cytosol. J Biol Chem. 1994;269(30):19321–19330. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
