

Notch signaling regulates cardiomyocyte proliferation during zebrafish heart regeneration
- PMID: 24474765
- PMCID: PMC3910613
- DOI: 10.1073/pnas.1311705111
Notch signaling regulates cardiomyocyte proliferation during zebrafish heart regeneration
Abstract
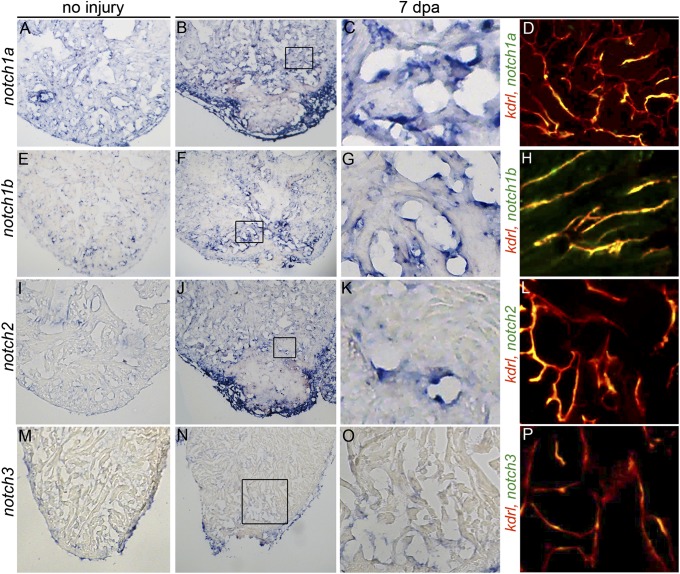
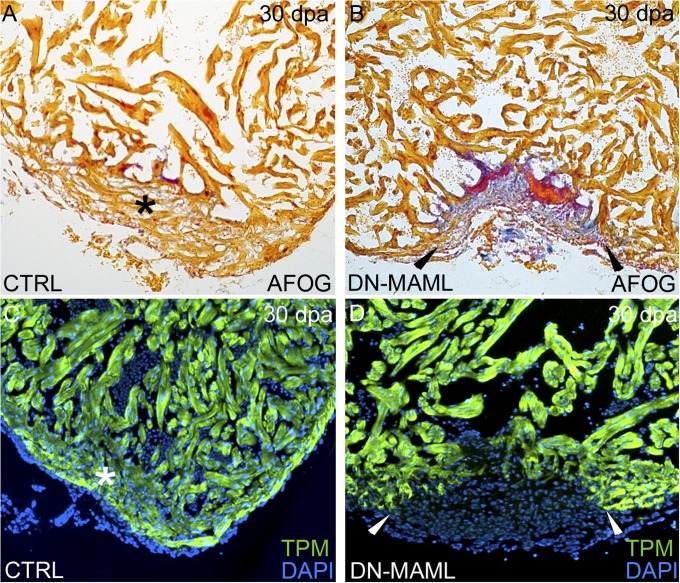
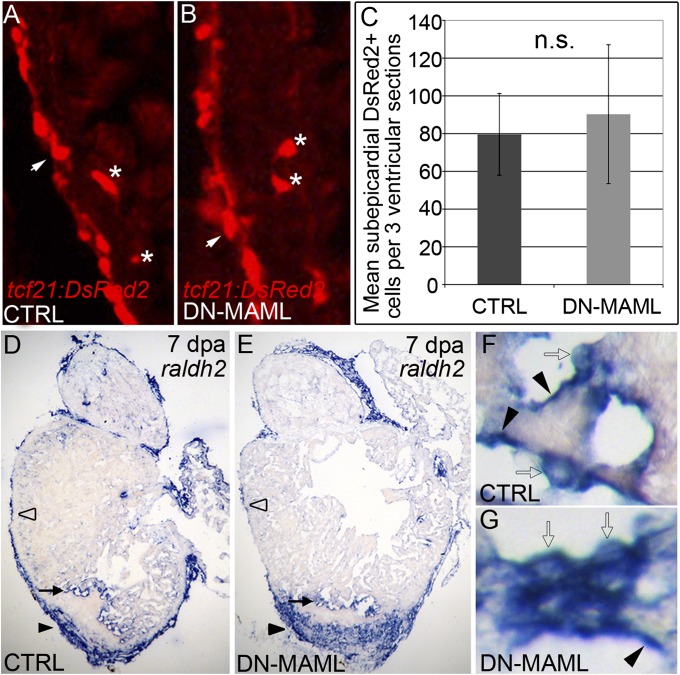
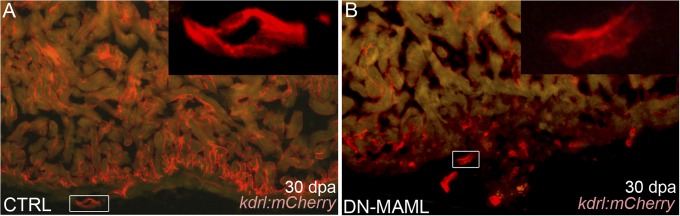
The human heart's failure to replace ischemia-damaged myocardium with regenerated muscle contributes significantly to the worldwide morbidity and mortality associated with coronary artery disease. Remarkably, certain vertebrate species, including the zebrafish, achieve complete regeneration of amputated or injured myocardium through the proliferation of spared cardiomyocytes. Nonetheless, the genetic and cellular determinants of natural cardiac regeneration remain incompletely characterized. Here, we report that cardiac regeneration in zebrafish relies on Notch signaling. Following amputation of the zebrafish ventricular apex, Notch receptor expression becomes activated specifically in the endocardium and epicardium, but not the myocardium. Using a dominant negative approach, we discovered that suppression of Notch signaling profoundly impairs cardiac regeneration and induces scar formation at the amputation site. We ruled out defects in endocardial activation, epicardial activation, and dedifferentiation of compact myocardial cells as causative for the regenerative failure. Furthermore, coronary endothelial tubes, which we lineage traced from preexisting endothelium in wild-type hearts, formed in the wound despite the myocardial regenerative failure. Quantification of myocardial proliferation in Notch-suppressed hearts revealed a significant decrease in cycling cardiomyocytes, an observation consistent with a noncell autonomous requirement for Notch signaling in cardiomyocyte proliferation. Unexpectedly, hyperactivation of Notch signaling also suppressed cardiomyocyte proliferation and heart regeneration. Taken together, our data uncover the exquisite sensitivity of regenerative cardiomyocyte proliferation to perturbations in Notch signaling.
Keywords: model organism; myocardial infarction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
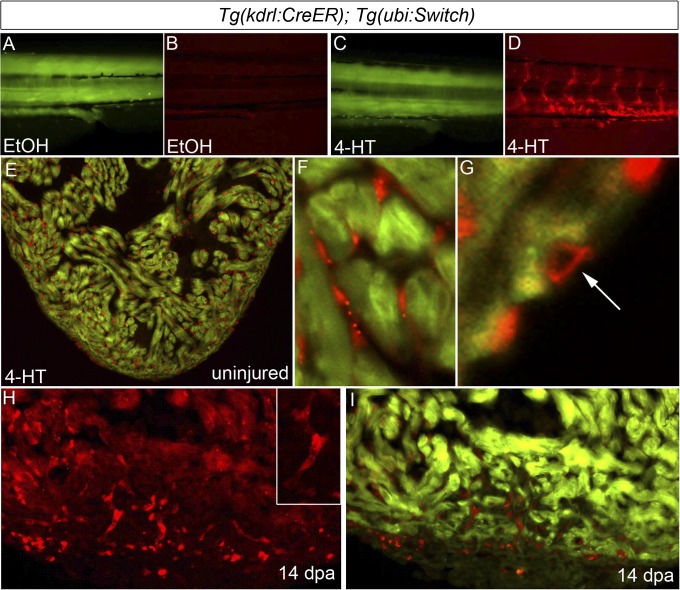
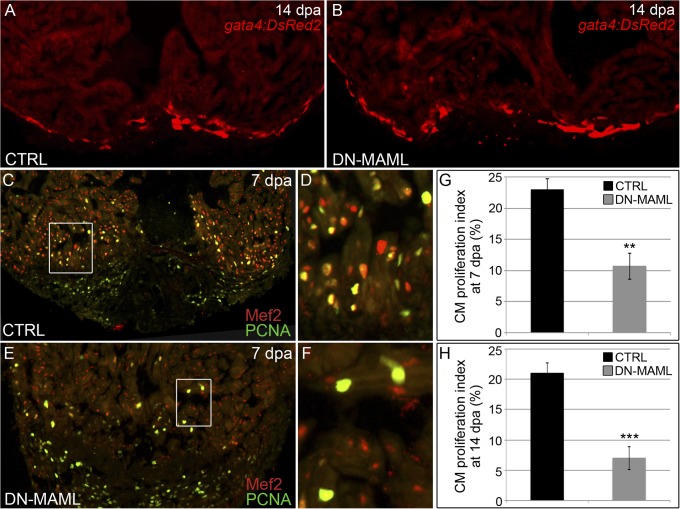
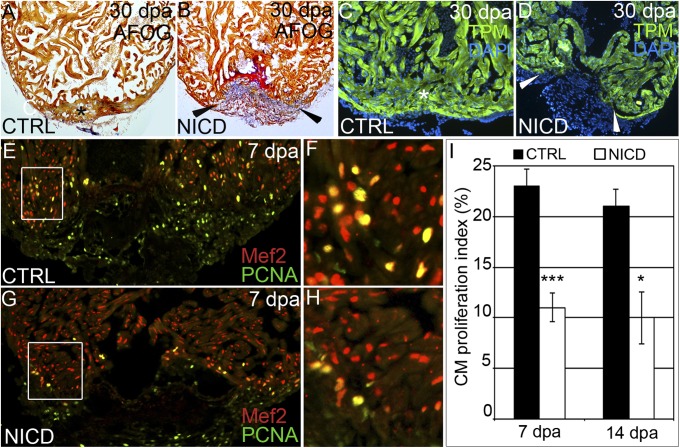
References
-
- Sidney S, Rosamond WD, Howard VJ, Luepker RV. National Forum for Heart Disease and Stroke Prevention The “heart disease and stroke statistics—2013 update” and the need for a national cardiovascular surveillance system. Circulation. 2013;127(1):21–23. - PubMed
-
- González-Rosa JM, Martín V, Peralta M, Torres M, Mercader N. Extensive scar formation and regression during heart regeneration after cryoinjury in zebrafish. Development. 2011;138(9):1663–1674. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
