

Proteolytic cleavage of Ser52Pro variant transthyretin triggers its amyloid fibrillogenesis
- PMID: 24474780
- PMCID: PMC3910611
- DOI: 10.1073/pnas.1317488111
Proteolytic cleavage of Ser52Pro variant transthyretin triggers its amyloid fibrillogenesis
Abstract
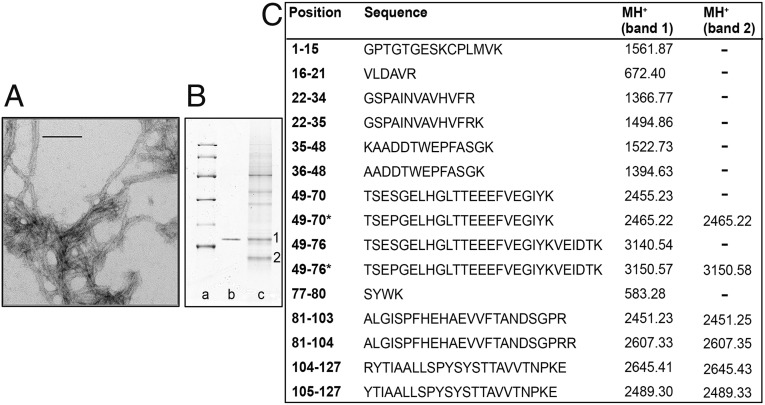
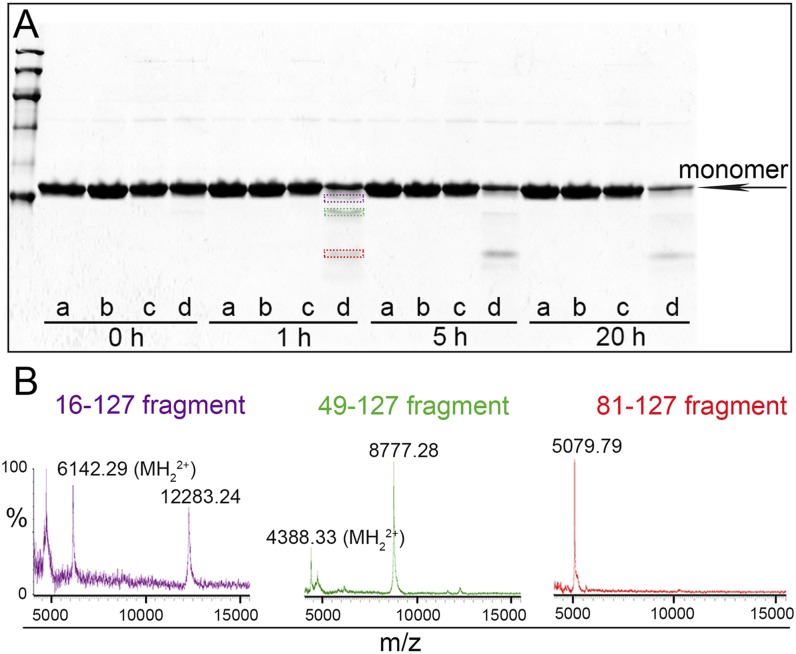
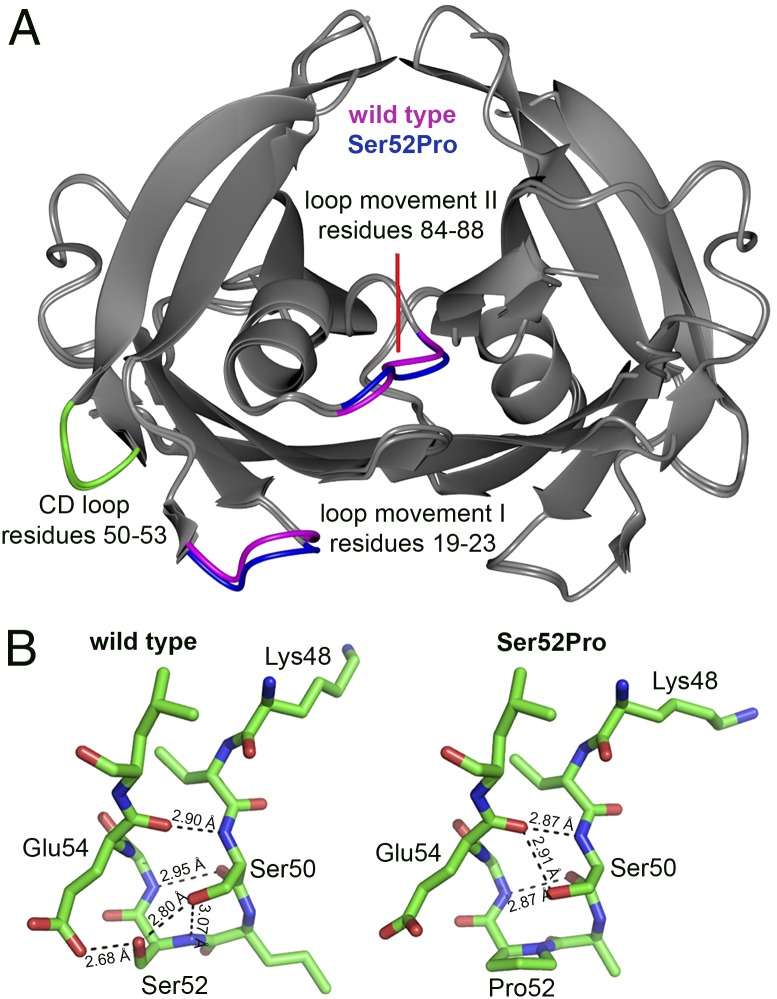
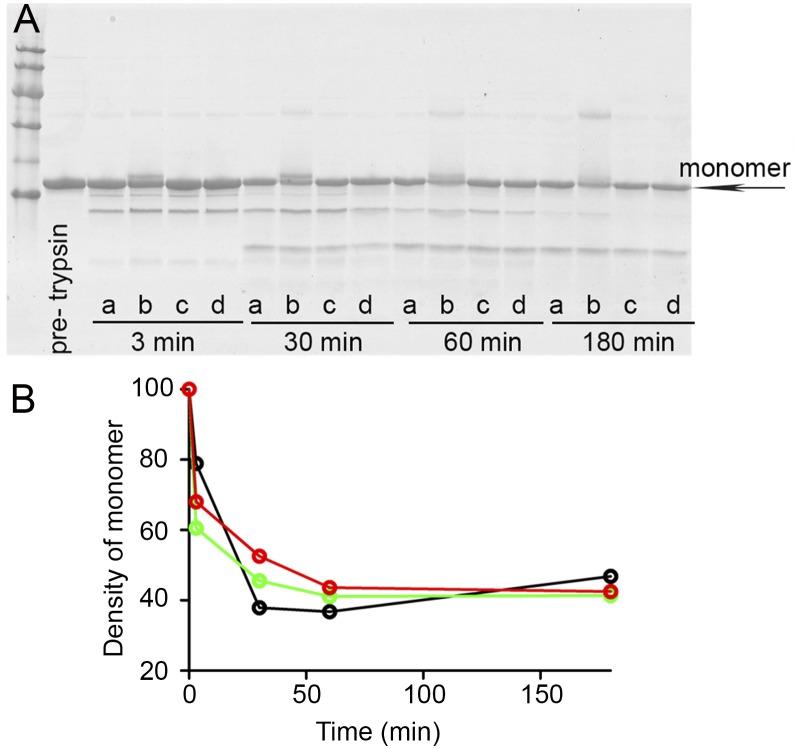
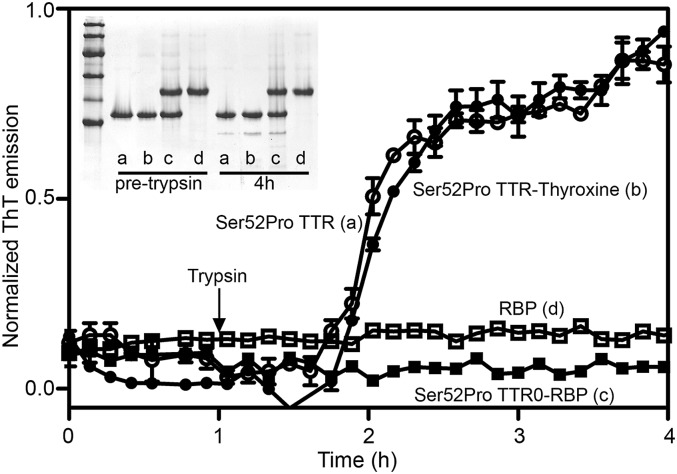
The Ser52Pro variant of transthyretin (TTR) produces aggressive, highly penetrant, autosomal-dominant systemic amyloidosis in persons heterozygous for the causative mutation. Together with a minor quantity of full-length wild-type and variant TTR, the main component of the ex vivo fibrils was the residue 49-127 fragment of the TTR variant, the portion of the TTR sequence that previously has been reported to be the principal constituent of type A, cardiac amyloid fibrils formed from wild-type TTR and other TTR variants [Bergstrom J, et al. (2005) J Pathol 206(2):224-232]. This specific truncation of Ser52Pro TTR was generated readily in vitro by limited proteolysis. In physiological conditions and under agitation the residue 49-127 proteolytic fragment rapidly and completely self-aggregates into typical amyloid fibrils. The remarkable susceptibility to such cleavage is likely caused by localized destabilization of the β-turn linking strands C and D caused by loss of the wild-type hydrogen-bonding network between the side chains of residues Ser52, Glu54, Ser50, and a water molecule, as revealed by the high-resolution crystallographic structure of Ser52Pro TTR. We thus provide a structural basis for the recently hypothesized, crucial pathogenic role of proteolytic cleavage in TTR amyloid fibrillogenesis. Binding of the natural ligands thyroxine or retinol-binding protein (RBP) by Ser52Pro variant TTR stabilizes the native tetrameric assembly, but neither protected the variant from proteolysis. However, binding of RBP, but not thyroxine, inhibited subsequent fibrillogenesis.
Keywords: misfolding; protein aggregation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Pepys MB. Amyloidosis. Annu Rev Med. 2006;57(1):223–241. - PubMed
-
- Merlini G, Bellotti V. Molecular mechanisms of amyloidosis. N Engl J Med. 2003;349(6):583–596. - PubMed
-
- Valleix S, et al. Hereditary systemic amyloidosis due to Asp76Asn variant β2-microglobulin. N Engl J Med. 2012;366(24):2276–2283. - PubMed
-
- Palaninathan SK. Nearly 200 X-ray crystal structures of transthyretin: What do they tell us about this protein and the design of drugs for TTR amyloidoses? Curr Med Chem. 2012;19(15):2324–2342. - PubMed
-
- Hammarström P, Wiseman RL, Powers ET, Kelly JW. Prevention of transthyretin amyloid disease by changing protein misfolding energetics. Science. 2003;299(5607):713–716. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
