

Structures of yeast mitochondrial ADP/ATP carriers support a domain-based alternating-access transport mechanism
- PMID: 24474793
- PMCID: PMC3910652
- DOI: 10.1073/pnas.1320692111
Structures of yeast mitochondrial ADP/ATP carriers support a domain-based alternating-access transport mechanism
Abstract
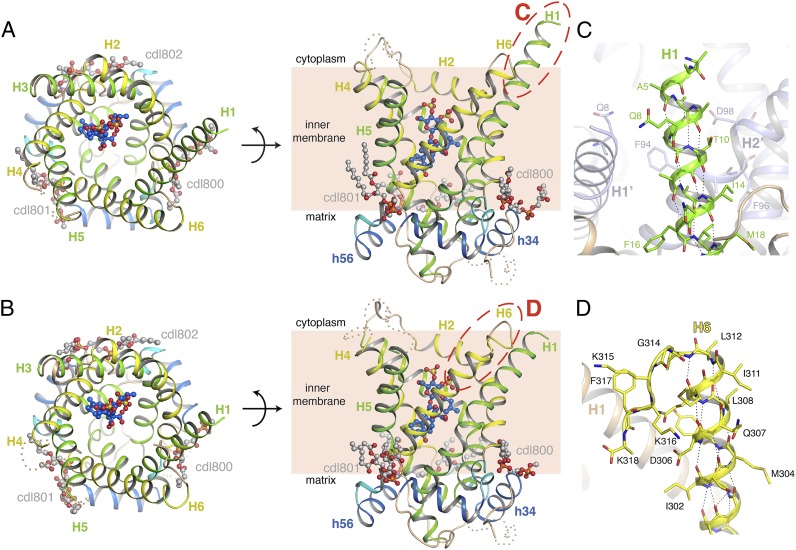
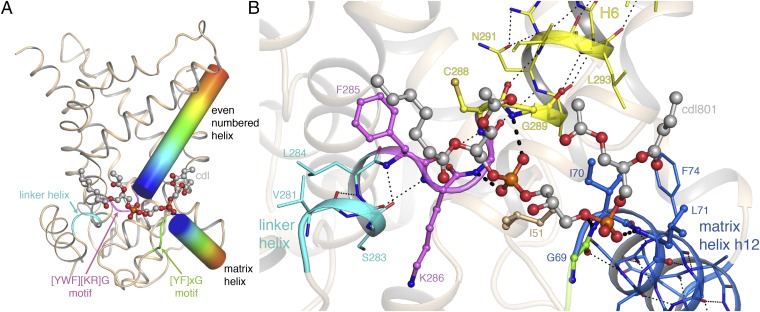
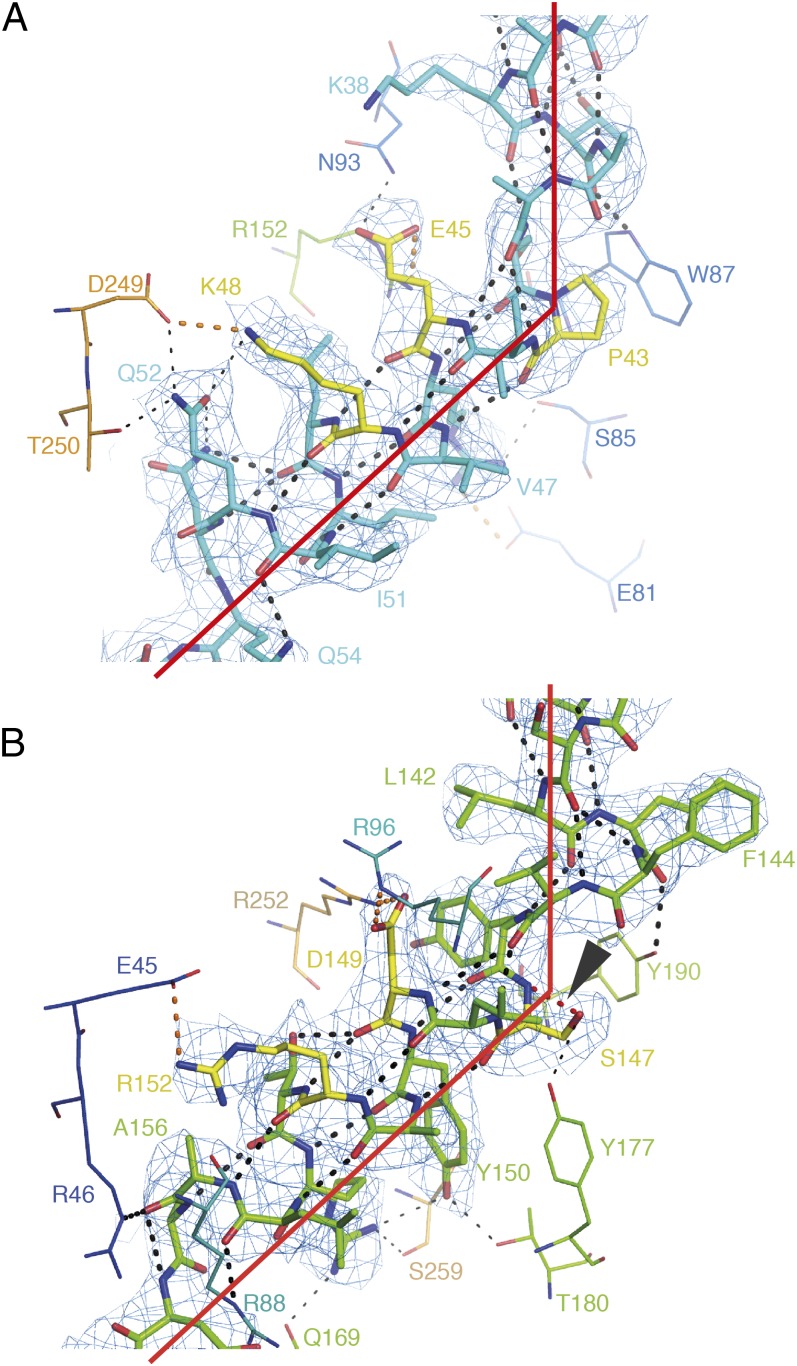
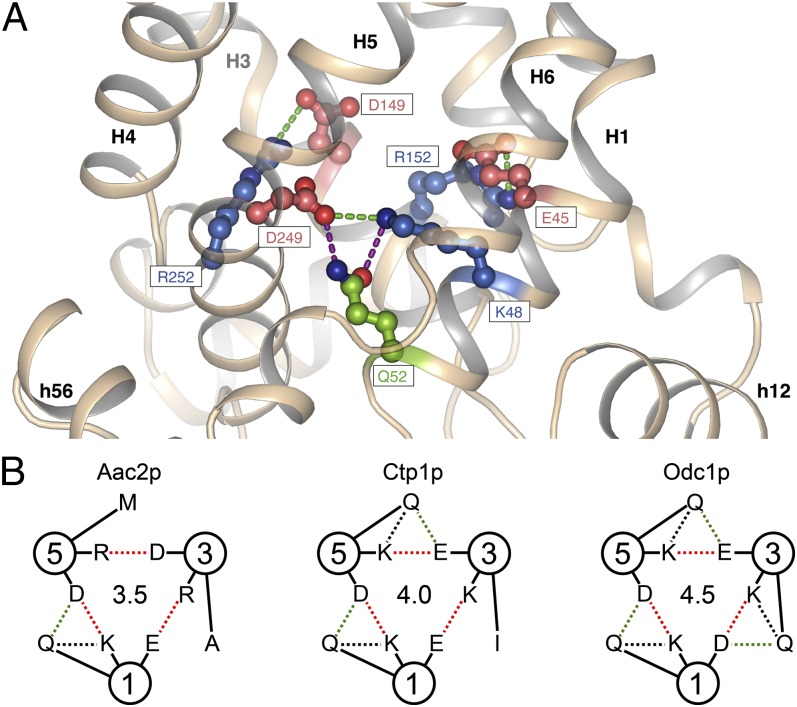
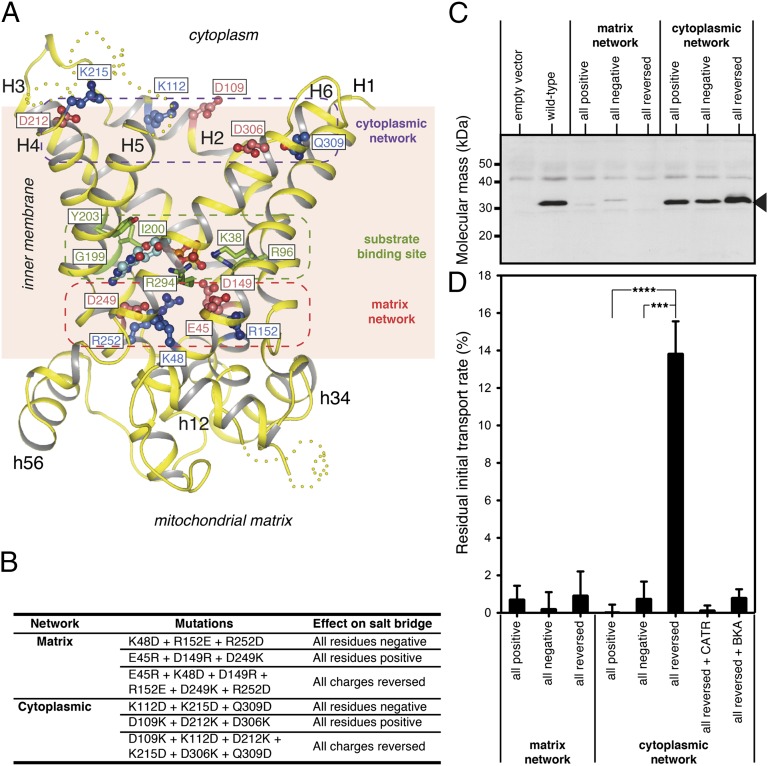
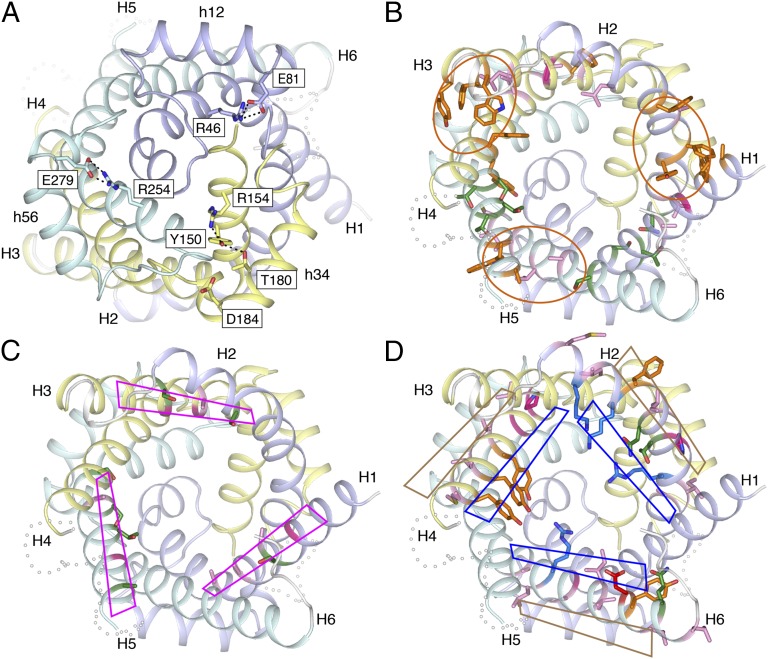
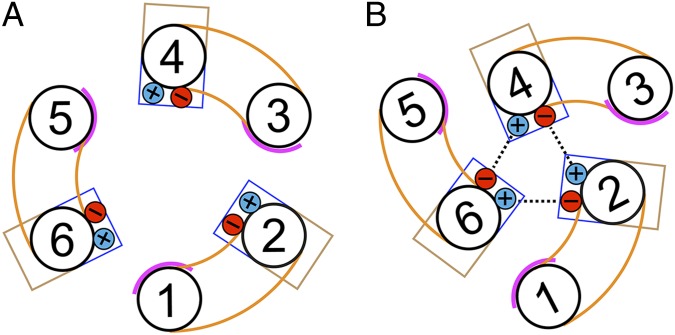
The mitochondrial ADP/ATP carrier imports ADP from the cytosol and exports ATP from the mitochondrial matrix. The carrier cycles by an unresolved mechanism between the cytoplasmic state, in which the carrier accepts ADP from the cytoplasm, and the matrix state, in which it accepts ATP from the mitochondrial matrix. Here we present the structures of the yeast ADP/ATP carriers Aac2p and Aac3p in the cytoplasmic state. The carriers have three domains and are closed at the matrix side by three interdomain salt-bridge interactions, one of which is braced by a glutamine residue. Glutamine braces are conserved in mitochondrial carriers and contribute to an energy barrier, preventing the conversion to the matrix state unless substrate binding occurs. At the cytoplasmic side a second salt-bridge network forms during the transport cycle, as demonstrated by functional analysis of mutants with charge-reversed networks. Analyses of the domain structures and properties of the interdomain interfaces indicate that interconversion between states involves movement of the even-numbered α-helices across the surfaces of the odd-numbered α-helices by rotation of the domains. The odd-numbered α-helices have an L-shape, with proline or serine residues at the kinks, which functions as a lever-arm, coupling the substrate-induced disruption of the matrix network to the formation of the cytoplasmic network. The simultaneous movement of three domains around a central translocation pathway constitutes a unique mechanism among transport proteins. These findings provide a structural description of transport by mitochondrial carrier proteins, consistent with an alternating-access mechanism.
Keywords: X-ray crystallography; adenine nucleotide translocase; cardiolipin binding; membrane protein; serine kinks.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Palmieri F. The mitochondrial transporter family SLC25: Identification, properties and physiopathology. Mol Aspects Med. 2013;34(2–3):465–484. - PubMed
-
- Palmieri F. Diseases caused by defects of mitochondrial carriers: A review. Biochim Biophys Acta. 2008;1777(7–8):564–578. - PubMed
-
- Klingenberg M. The ADP and ATP transport in mitochondria and its carrier. Biochim Biophys Acta. 2008;1778(10):1978–2021. - PubMed
-
- Duee ED, Vignais PV. [Exchange between extra- and intramitochondrial adenine nucleotides] Biochim Biophys Acta. 1965;107(1):184–188. French. - PubMed
-
- Vignais PV, Vignais PM, Defaye G. Adenosine diphosphate translocation in mitochondria. Nature of the receptor site for carboxyatractyloside (gummiferin) Biochemistry. 1973;12(8):1508–1519. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
