

S-palmitoylation regulates biogenesis of core glycosylated wild-type and F508del CFTR in a post-ER compartment
- PMID: 24475974
- PMCID: PMC3993085
- DOI: 10.1042/BJ20131037
S-palmitoylation regulates biogenesis of core glycosylated wild-type and F508del CFTR in a post-ER compartment
Abstract
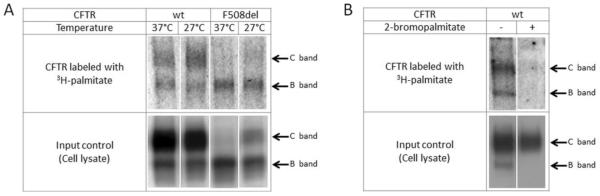
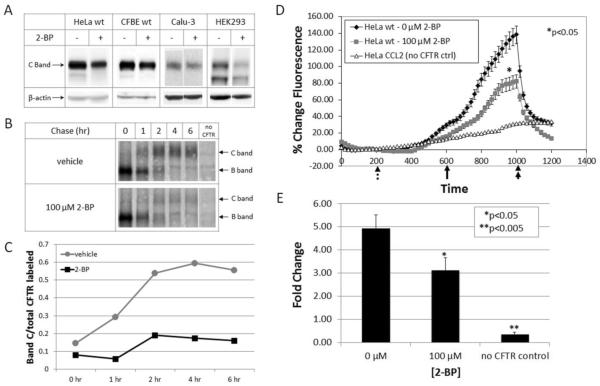
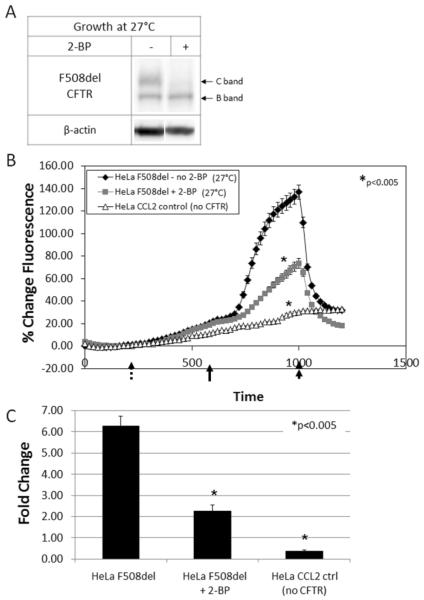
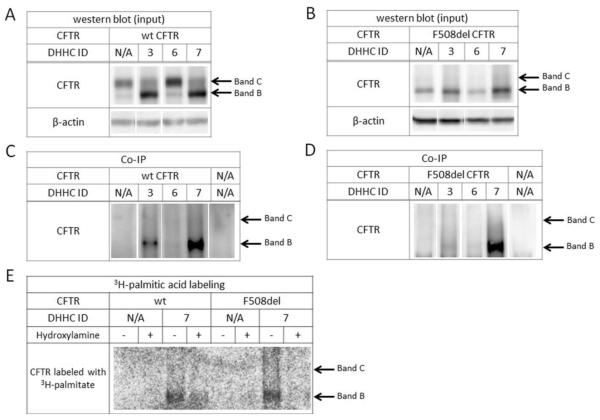
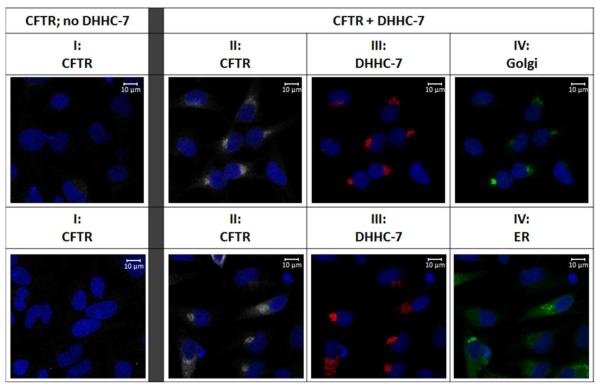
Defects in CFTR (cystic fibrosis transmembrane conductance regulator) maturation are central to the pathogenesis of CF (cystic fibrosis). Palmitoylation serves as a key regulator of maturational processing in other integral membrane proteins, but has not been tested previously for functional effects on CFTR. In the present study, we used metabolic labelling to confirm that wild-type and F508del CFTR are palmitoylated, and show that blocking palmitoylation with the pharmacologic inhibitor 2-BP (2-bromopalmitate) decreases steady-state levels of both wild-type and low temperature-corrected F508del CFTR, disrupts post-ER (endoplasmic reticulum) maturation and reduces ion channel function at the cell surface. PATs (protein acyl transferases) comprise a family of 23 gene products that contain a DHHC motif and mediate palmitoylation. Recombinant expression of specific PATs led to increased levels of CFTR protein and enhanced palmitoylation as judged by Western blot and metabolic labelling. Specifically, we show that DHHC-7 (i) increases steady-state levels of wild-type and F508del CFTR band B, (ii) interacts preferentially with the band B glycoform, and (iii) augments radiolabelling by [3H]palmitic acid. Interestingly, immunofluorescence revealed that DHHC-7 also sequesters the F508del protein to a post-ER (Golgi) compartment. Our findings point to the importance of palmitoylation during wild-type and F508del CFTR trafficking.
Figures
References
-
- Ward CL, Omura S, Kopito RR. Degradation of CFTR by the ubiquitin-proteasome pathway. Cell. 1995;83:121–127. - PubMed
-
- Gelman MS, Kannegaard ES, Kopito RR. A principal role for the proteasome in endoplasmic reticulum-associated degradation of misfolded intracellular cystic fibrosis transmembrane conductance regulator. J. Biol. Chem. 2002;277:11709–11714. - PubMed
-
- Swiatecka-Urban A, Brown A, Moreau-Marquis S, Renuka J, Coutermarsh B, Barnaby R, Karlson KH, Flotte TR, Fukuda M, Langford GM, Stanton BA. The short apical membrane half-life of rescued {Delta}F508-cystic fibrosis transmembrane conductance regulator (CFTR) results from accelerated endocytosis of {Delta}F508-CFTR in polarized human airway epithelial cells. J. Biol. Chem. 2005;280:36762–36772. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources