

Temperature sensitivity of acid-sensitive outwardly rectifying (ASOR) anion channels in cortical neurons is involved in hypothermic neuroprotection against acidotoxic necrosis
- PMID: 24476793
- PMCID: PMC4203757
- DOI: 10.4161/chan.27748
Temperature sensitivity of acid-sensitive outwardly rectifying (ASOR) anion channels in cortical neurons is involved in hypothermic neuroprotection against acidotoxic necrosis
Abstract
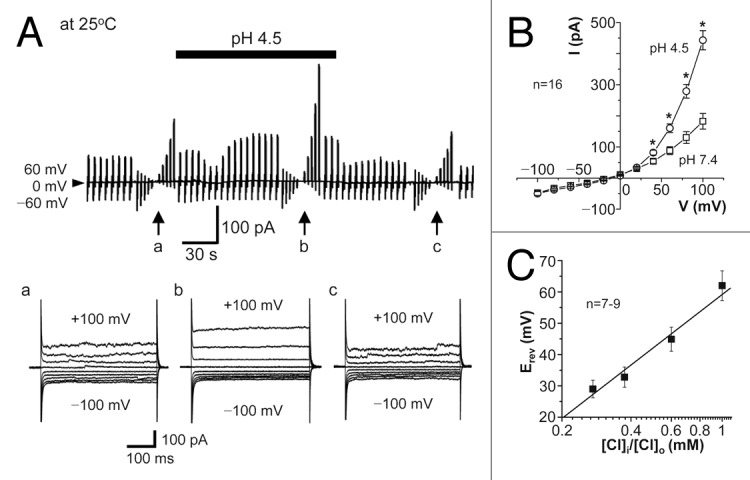
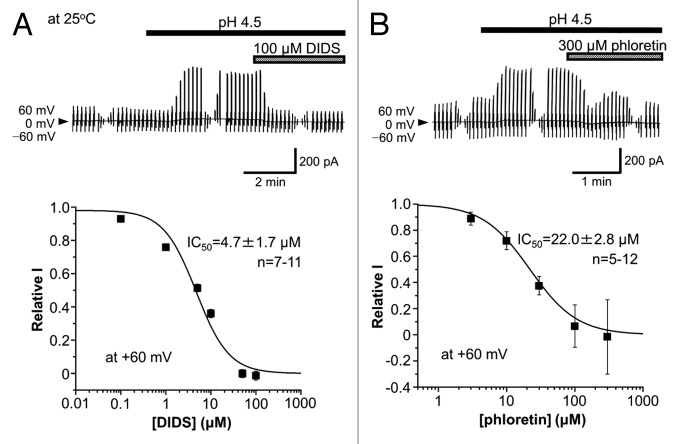
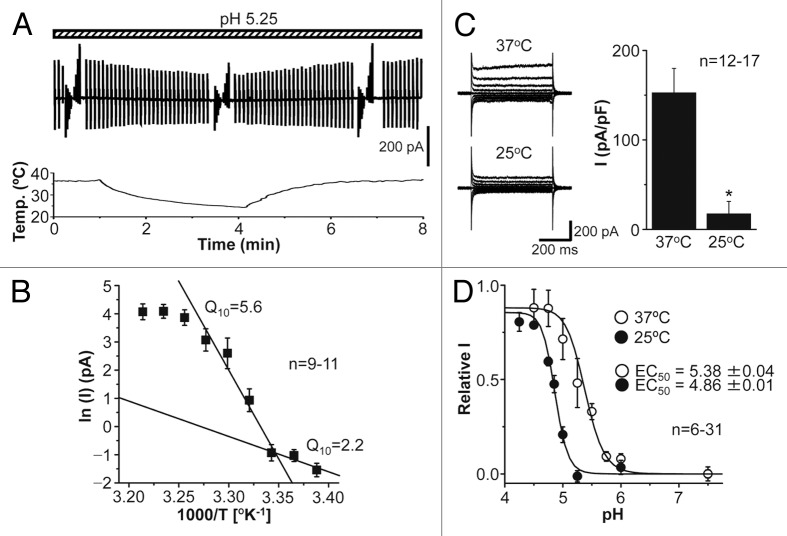
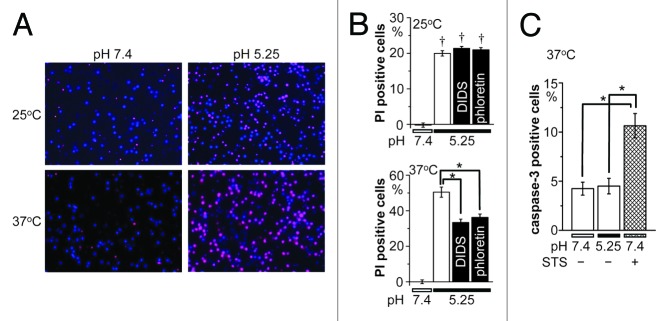
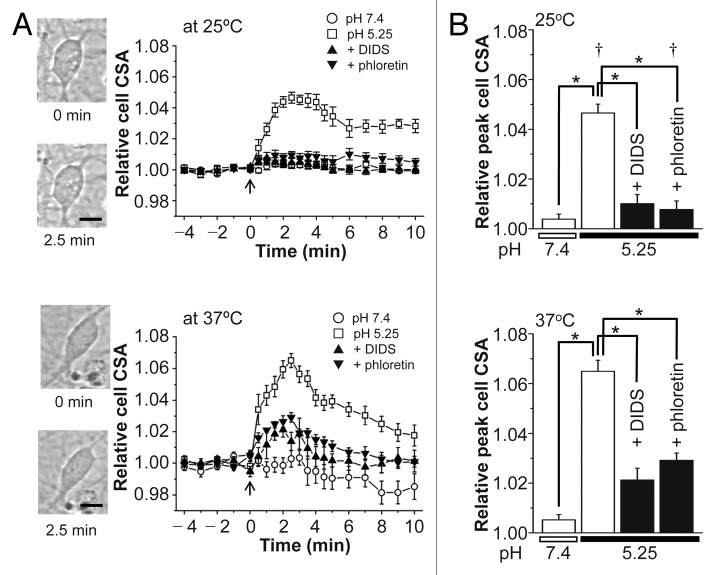
The acid-sensitive outwardly rectifying (ASOR) anion channel has been found in non-neuronal cell types and was shown to be involved in acidotoxic death of epithelial cells. We have recently shown that the ASOR channel is sensitive to temperature. Here, we extend those results to show that temperature-sensitive ASOR anion channels are expressed in cortical neurons and involved in acidotoxic neuronal cell death. In cultured mouse cortical neurons, reduction of extracellular pH activated anionic currents exhibiting phenotypic properties of the ASOR anion channel. The neuronal ASOR currents recorded at pH 5.25 were augmented by warm temperature, with a threshold temperature of 26 °C and the Q(10) value of 5.6. After 1 h exposure to acidic solution at 37 °C, a large population of neurons suffered from necrotic cell death which was largely protected not only by ASOR channel blockers but also by reduction of temperature to 25 °C. Thus, it is suggested that high temperature sensitivity of the neuronal ASOR anion channel provides, at least in part, a basis for hypothermic neuroprotection under acidotoxic situations associated with a number of pathological brain states.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical