

The α isoform of topoisomerase II is required for hypercompaction of mitotic chromosomes in human cells
- PMID: 24476913
- PMCID: PMC3985649
- DOI: 10.1093/nar/gku076
The α isoform of topoisomerase II is required for hypercompaction of mitotic chromosomes in human cells
Abstract
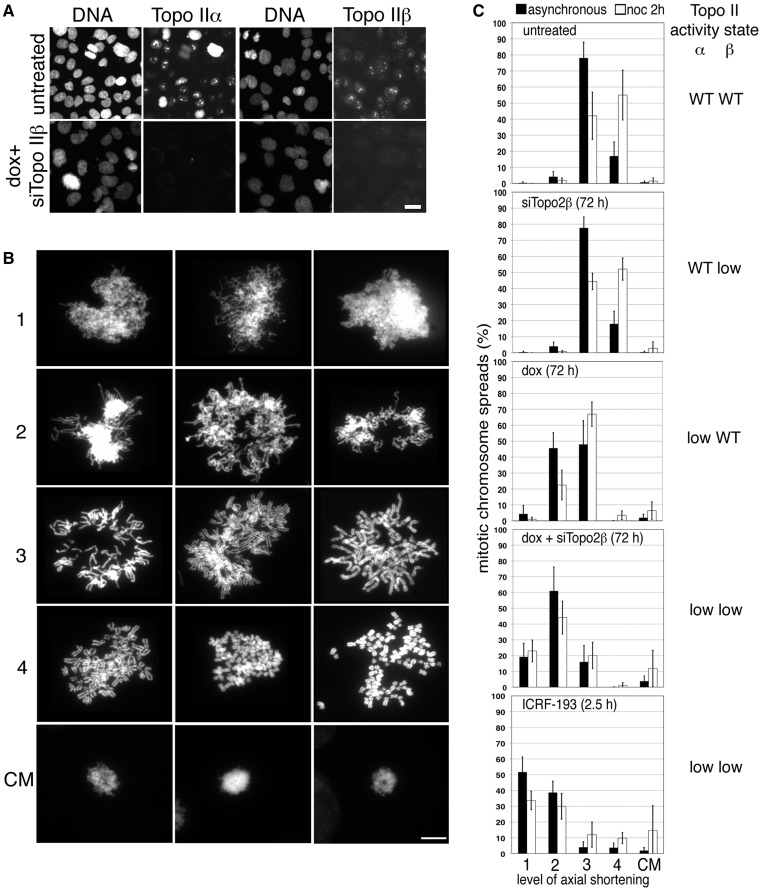
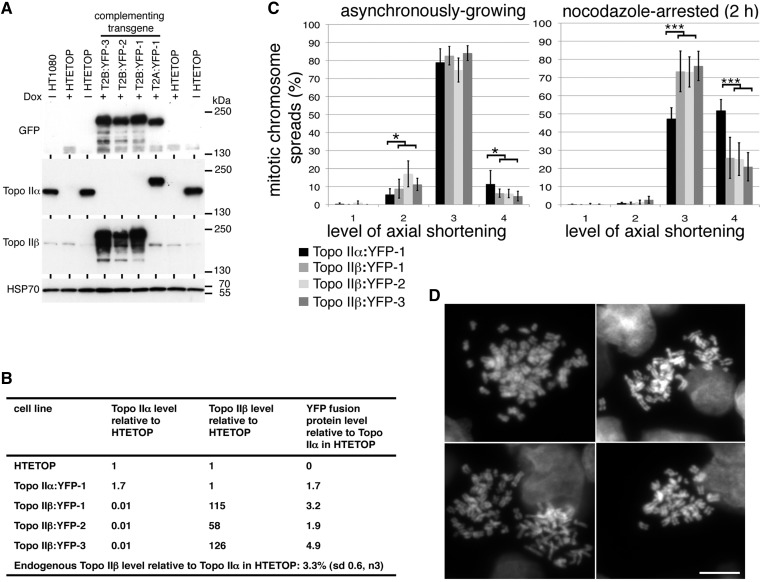
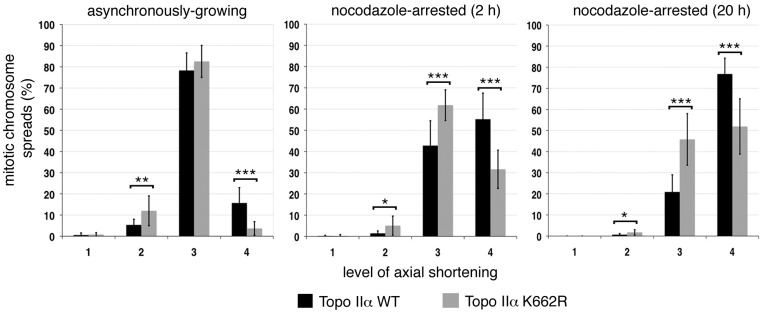
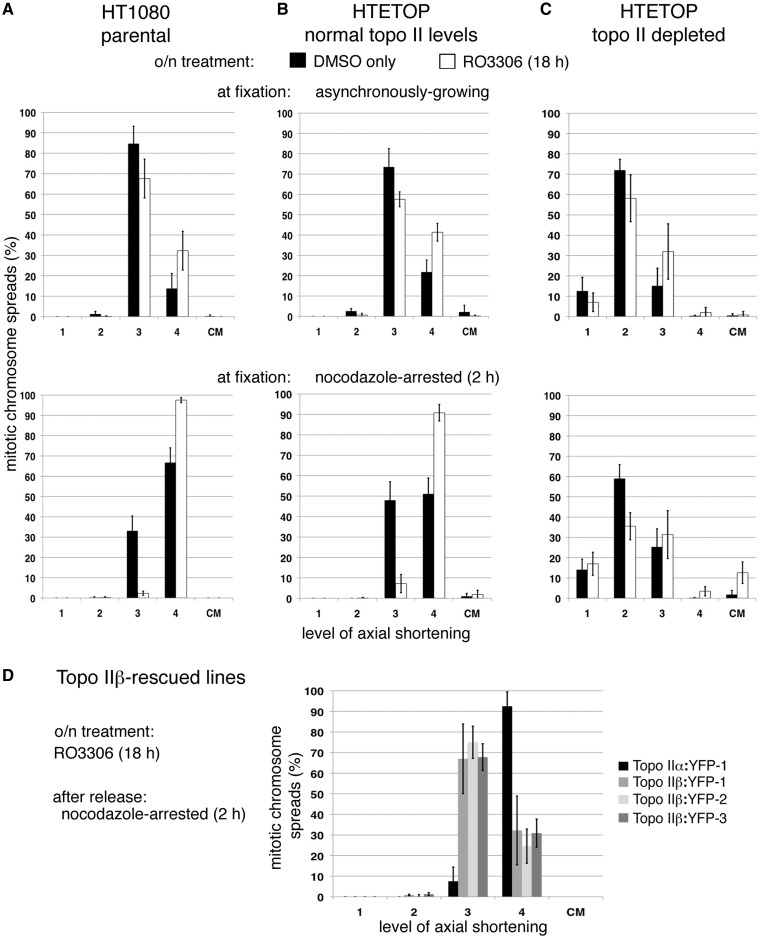
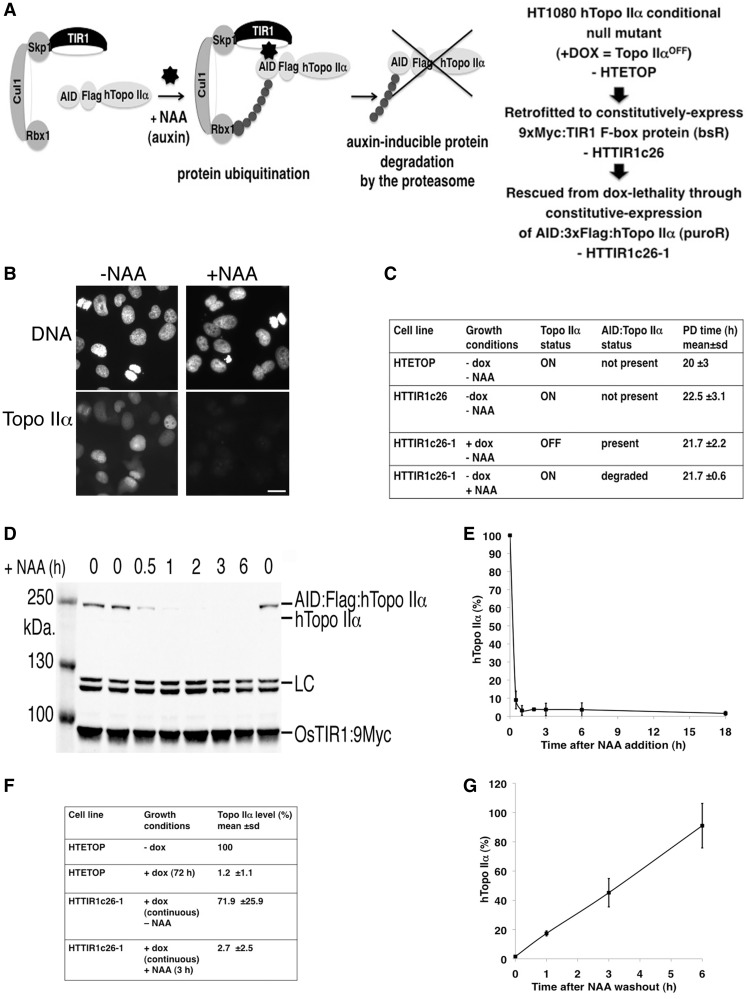
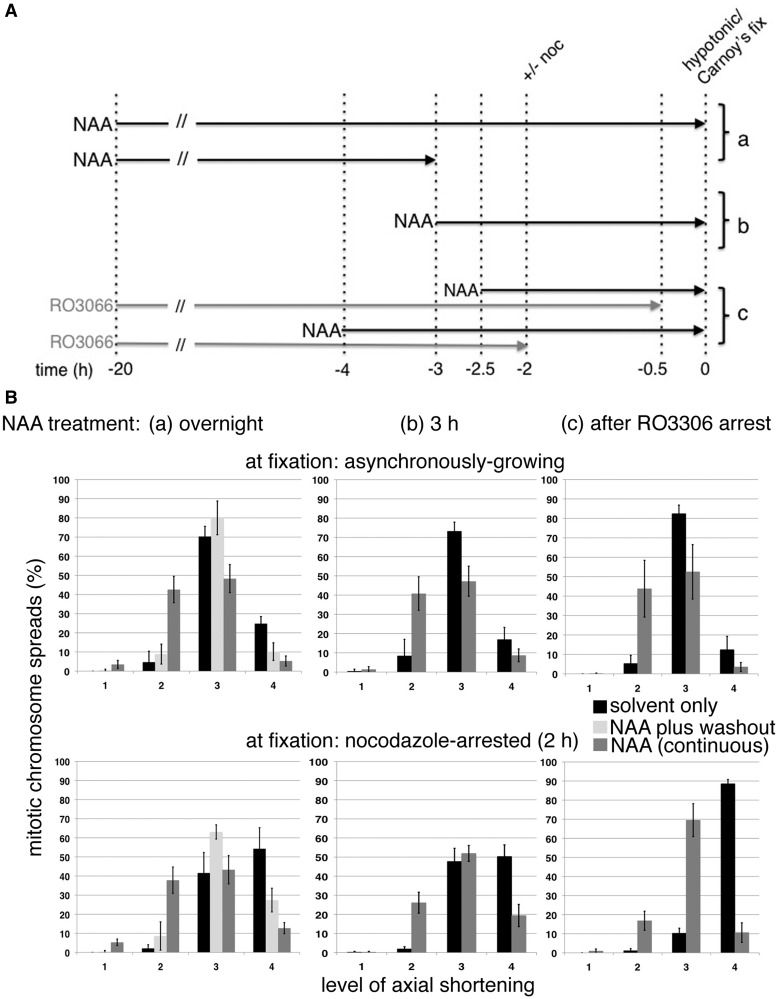
As proliferating cells transit from interphase into M-phase, chromatin undergoes extensive reorganization, and topoisomerase (topo) IIα, the major isoform of this enzyme present in cycling vertebrate cells, plays a key role in this process. In this study, a human cell line conditional null mutant for topo IIα and a derivative expressing an auxin-inducible degron (AID)-tagged version of the protein have been used to distinguish real mitotic chromosome functions of topo IIα from its more general role in DNA metabolism and to investigate whether topo IIβ makes any contribution to mitotic chromosome formation. We show that topo IIβ does contribute, with endogenous levels being sufficient for the initial stages of axial shortening. However, a significant effect of topo IIα depletion, seen with or without the co-depletion of topo IIβ, is the failure of chromosomes to hypercompact when delayed in M-phase. This requires much higher levels of topo II protein and is impaired by drugs or mutations that affect enzyme activity. A prolonged delay at the G2/M border results in hyperefficient axial shortening, a process that is topo IIα-dependent. Rapid depletion of topo IIα has allowed us to show that its function during late G2 and M-phase is truly required for shaping mitotic chromosomes.
Figures
References
-
- Tan KB, Dorman TE, Falls KM, Chung TD, Mirabelli CK, Crooke ST, Mao J. Topoisomerase II alpha and topoisomerase II beta genes: characterization and mapping to human chromosomes 17 and 3, respectively. Cancer Res. 1992;52:231–234. - PubMed
-
- Niimi A, Suka N, Harata M, Kikuchi A, Mizuno S. Co-localization of chicken DNA topoisomerase IIalpha, but not beta, with sites of DNA replication and possible involvement of a C-terminal region of alpha through its binding to PCNA. Chromosoma. 2001;110:102–114. - PubMed
-
- Adachi N, Nomoto M, Kohno K, Koyama H. Cell-cycle regulation of the DNA topoisomerase IIalpha promoter is mediated by proximal CCAAT boxes: possible involvement of acetylation. Gene. 2000;245:49–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
