

Review
doi: 10.3389/fnbeh.2013.00224.
eCollection 2013.
Sleep and protein synthesis-dependent synaptic plasticity: impacts of sleep loss and stress
Affiliations
- PMID: 24478645
- PMCID: PMC3896837
- DOI: 10.3389/fnbeh.2013.00224
Item in Clipboard
Review
Sleep and protein synthesis-dependent synaptic plasticity: impacts of sleep loss and stress
Front Behav Neurosci.
.
Abstract
Sleep has been ascribed a critical role in cognitive functioning. Several lines of evidence implicate sleep in the consolidation of synaptic plasticity and long-term memory. Stress disrupts sleep while impairing synaptic plasticity and cognitive performance. Here, we discuss evidence linking sleep to mechanisms of protein synthesis-dependent synaptic plasticity and synaptic scaling. We then consider how disruption of sleep by acute and chronic stress may impair these mechanisms and degrade sleep function.
Keywords: Arc/Arg3.1; brain-derived neurotrophic factor; gene expression; long-term potentiation; mood disorder; sleep deprivation; stress; translation control.
Figures
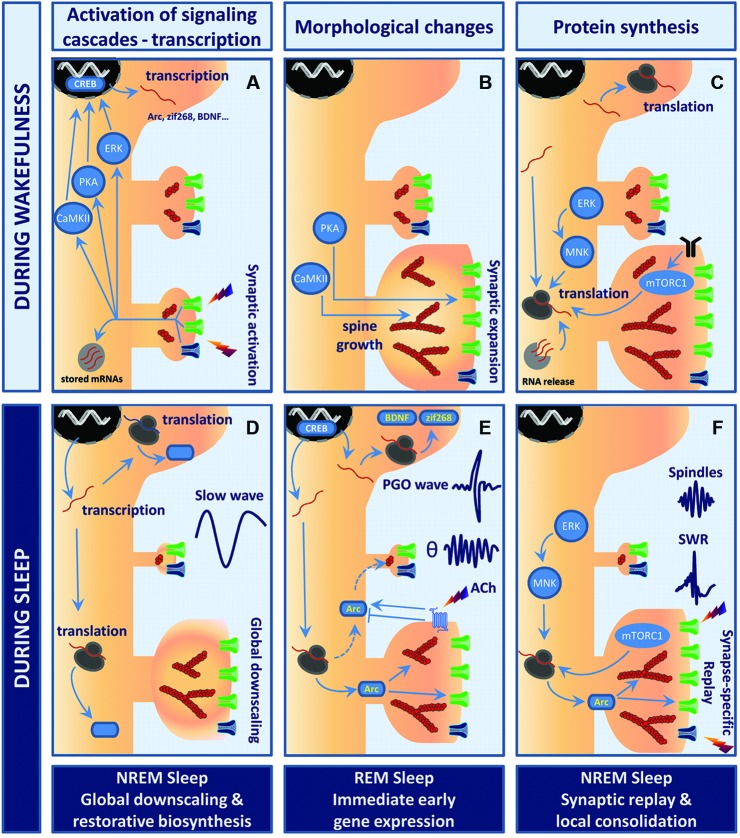
Model of sleep stage-specific potentiation and homeostatic scaling. In this working model, waking experience (LTP-like event) is consolidated through sleep stage-specific synaptic scaling, immediate early gene expression, and protein synthesis. Wakefulness (A–C). (A) Stimulation of glutamatergic synapses leads to rapid calcium influx into the postsynaptic compartment via NMDAR (dark blue) and AMPAR (green). Elevation of calcium levels activates multiple kinases and signaling cascades (e.g., PKA, CaMKII, ERK) which converge toward transcription factors such as cyclic AMP response element-binding protein (CREB), thus triggering rapid immediate early gene (IEG) expression. (B) Within minutes, polymerization of actin into filaments (red, actin filaments (F-actin)) induces remodeling of the actin cytoskeleton within spines. While CaMKII likely contributes to bundling of F-actin and expansion of the actin scaffolds, PKA promotes the insertion of AMPARs into the postsynaptic membrane. This whole process results in robust growth of synapses and enhanced synaptic efficacy. (C) These changes are then wake-consolidated in a de novo protein synthesis-dependent manner. Newly transcribed IEGs are either translated in the cell soma or trafficked further into the dendrites to be processed by the local translation machinery. Neuronal activity may release and translate dendritically stored mRNAs (light gray circle). Both mammalian target of rapamycin complex 1 (mTORC1) and mitogen-activated protein kinase-interacting kinase (MNK) signaling enhance rates of translation initiation. Postsynaptic receptors depicted are TrkB (black), AMPAR (green) and NMDAR (blue). Sleep (D–F). (D) NREM sleep supports the homeostatic process of cellular restoration by transcription and translation of genes involved in macromolecular biosynthesis and transport. In parallel, slow-wave activity (SWA) generates global synaptic downscaling in which synapses shrink and synaptic efficacy is reduced. Synapse-specific LTD at inactive or weakly active synapses may also be involved. (E) REM sleep and ponto-geniculo-occipital (PGO)-waves reactivate transcription of the plasticity-related IEGs Arc, brain-derived neurotrophic factor (BDNF) and zif268. Theta (θ) activity and increased acetylcholine levels regulate Arc protein turnover at the level of translation, degradation and mRNA decay. Arc may consolidate activity-induced synaptic changes by stabilizing the actin cytoskeleton and regulating trafficking of AMPAR from and to the postsynaptic membrane. (F) NREM sleep events like hippocampal sharp wave-ripples (SWRs) and thalamo-cortical sleep spindles have been suggested to actively take part in memory consolidation in conjunction with replay of neuronal activity patterns representing waking experience. The precise function of SWRs at the synaptic scale is yet to be unveiled. However, sparse and synapse-specific reactivations during SWRs of NREM sleep could provide bursts of local protein synthesis that consolidate synaptic modifications and memory formation. Thus, alternations of REM sleep-associated gene expression and NREM sleep-associated synaptic replay favor protein synthesis-dependent synaptic consolidation across sleep cycles.
References
-
- Artola A., von Frijtag J. C., Fermont P. C., Gispen W. H., Schrama L. H., Kamal A., et al. (2006). Long-lasting modulation of the induction of LTD and LTP in rat hippocampal CA1 by behavioural stress and environmental enrichment. Eur. J. Neurosci. 23, 261–272 10.1111/j.1460-9568.2005.04552.x - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical