

Concomitant inactivation of foxo3a and fancc or fancd2 reveals a two-tier protection from oxidative stress-induced hydrocephalus
- PMID: 24483844
- PMCID: PMC4186827
- DOI: 10.1089/ars.2013.5597
Concomitant inactivation of foxo3a and fancc or fancd2 reveals a two-tier protection from oxidative stress-induced hydrocephalus
Abstract
Aims: This study seeks at investigating the cause of hydrocephalus, and at identifying therapeutic targets for the prevention of hydrocephalus.
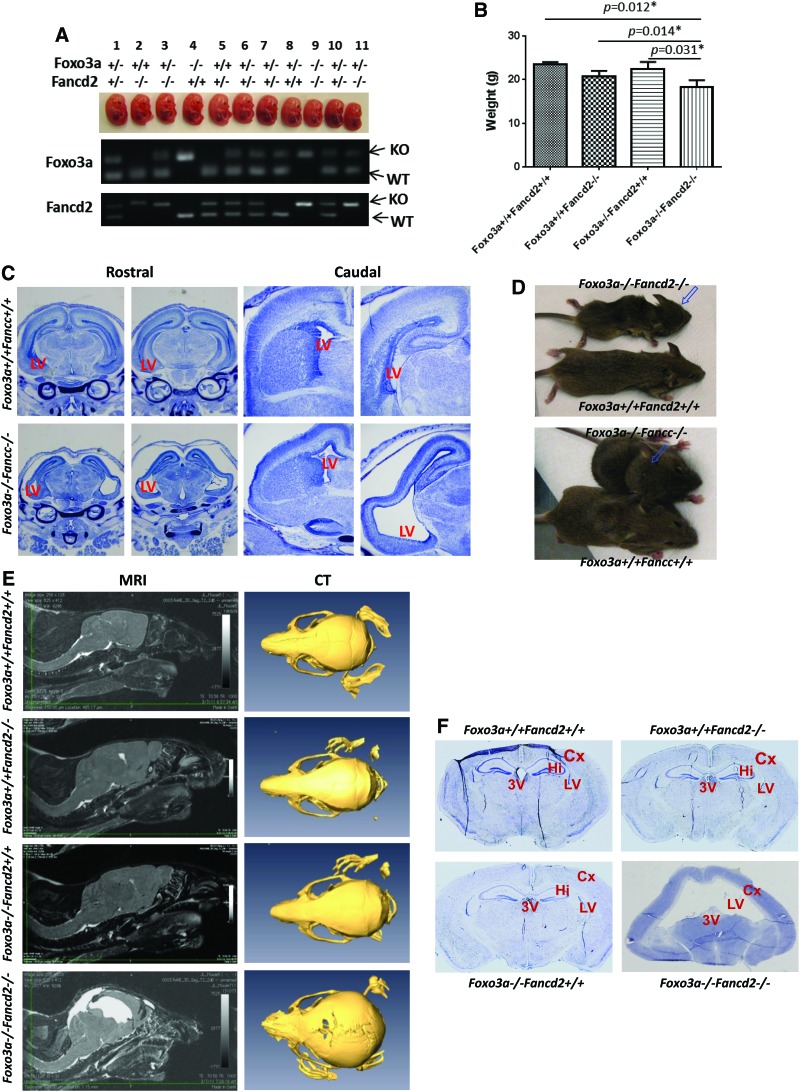
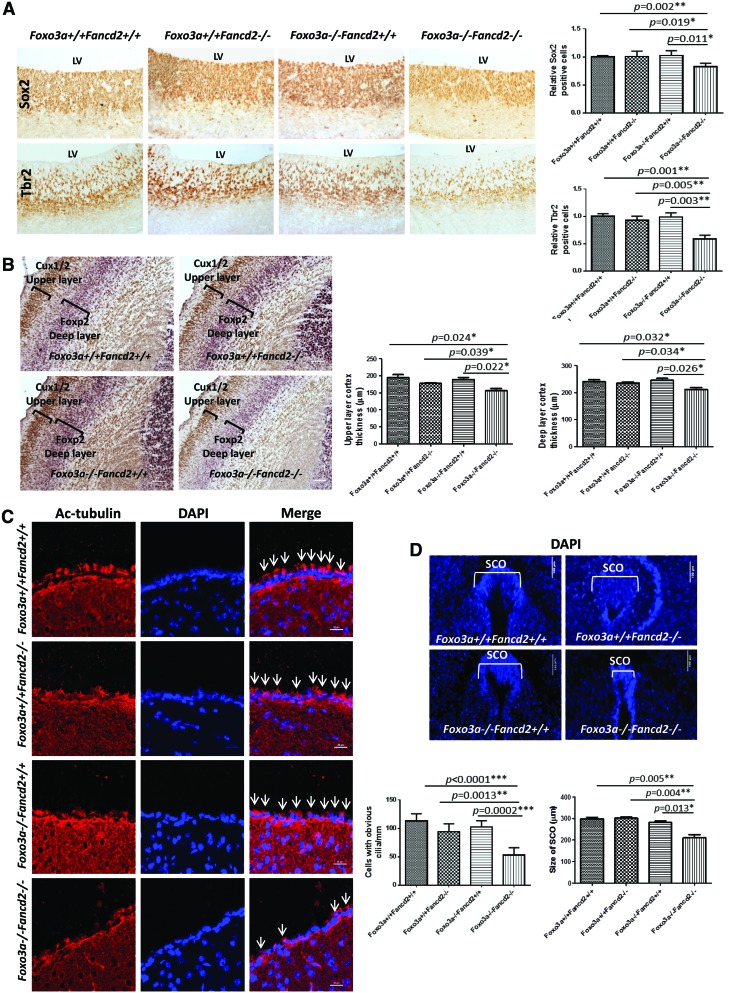
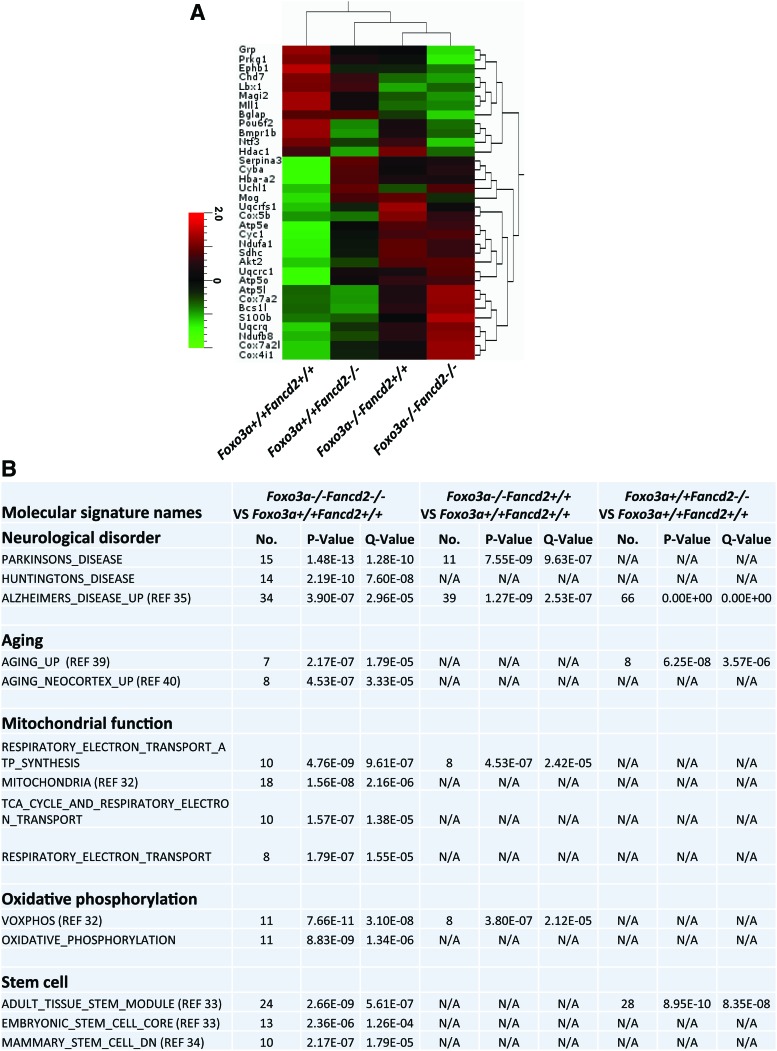
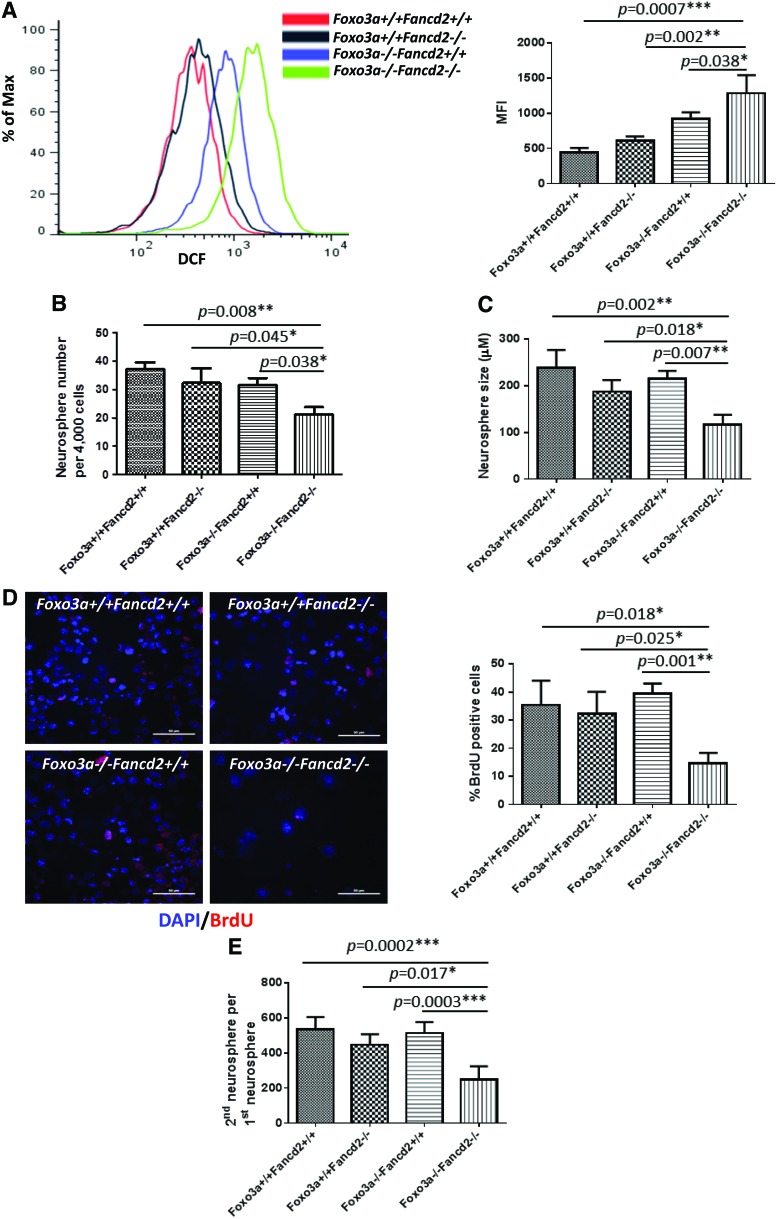
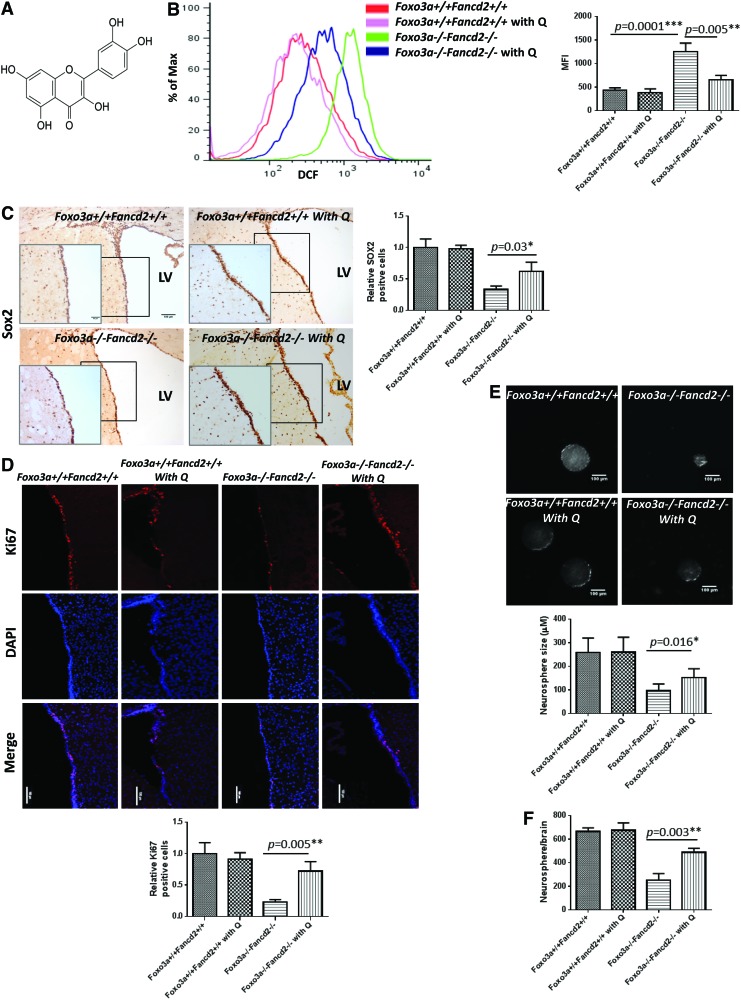
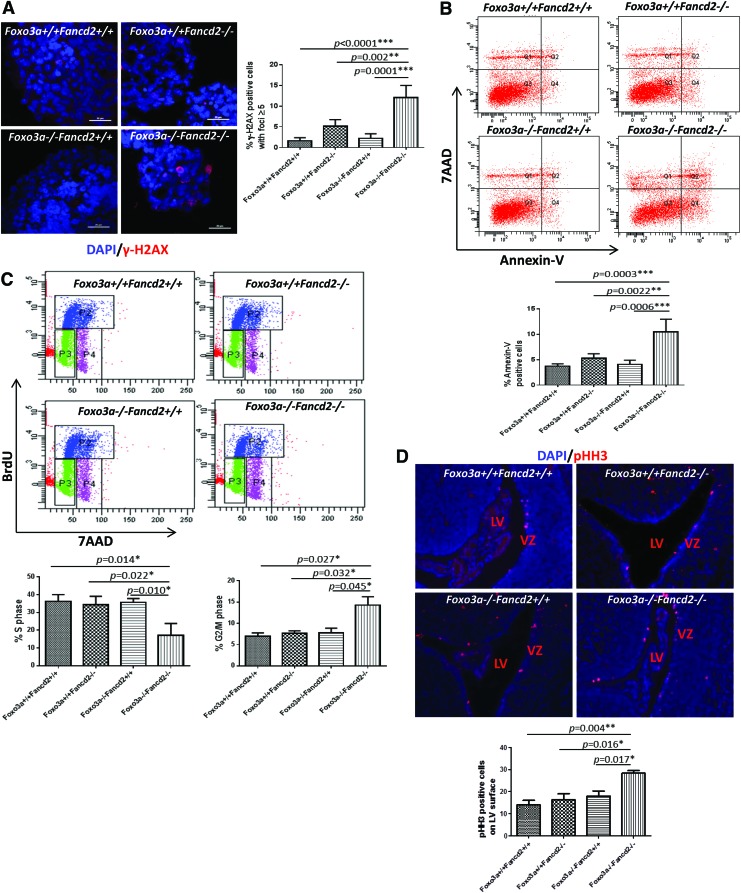
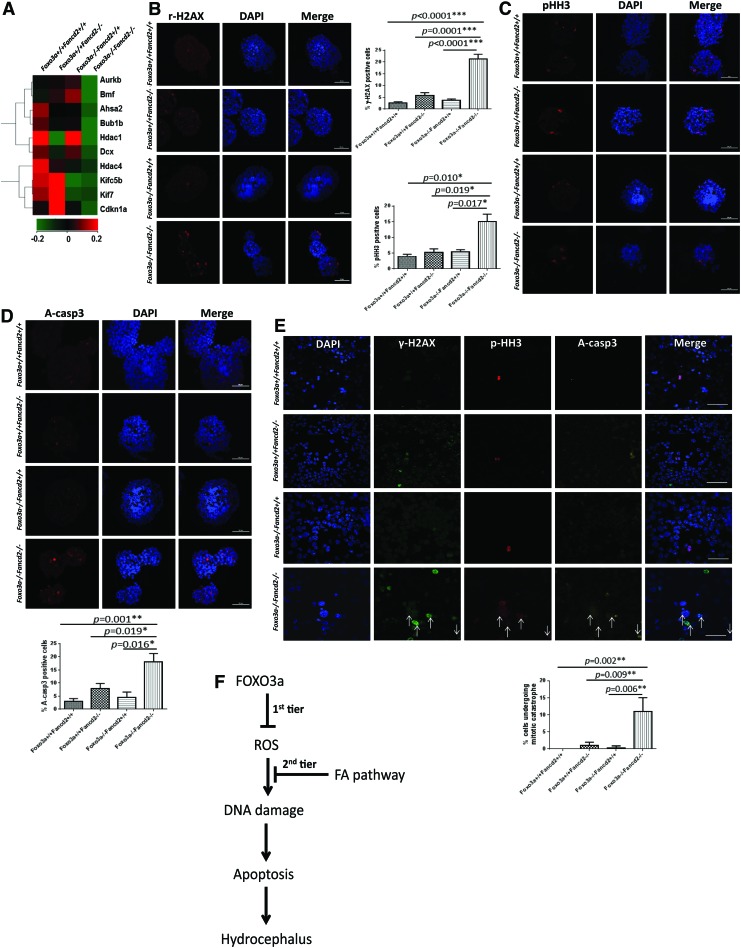
Results: In this study, we show that inactivation of the Foxo3a gene in two mouse models of Fanconi anemia (FA) leads to the development of hydrocephalus in late embryonic stage and after birth. More than 50% of Foxo3a(-/-) Fancc(-/-) or Foxo3a(-/-) Fancd2(-/-) mice die during embryonic development or within 6 months of life as a result of hydrocephalus characterized by cranial distortion, dilation of the ventricular system, reduced thickness of the cerebral cortex, and disorganization of the ependymal cilia and subcommissural organ. Combined deficiency of Foxo3a and Fancc or Fancd2 not only impairs the self-renewal capacity but also markedly increases the apoptosis of neural stem and progenitor cells (NSPCs), leading to defective neurogenesis. Increased accumulation of reactive oxygen species (ROS) and subsequently de-regulated mitosis and ultimately apoptosis in the neural stem or progenitor cells is identified as one of the potential mechanisms of congenital obstructive hydrocephalus.
Innovation: The work unravels a two-tier protective mechanism for preventing oxidative stress-induced hydrocephalus.
Conclusion: The deletion of Foxo3a in FA mice increased the accumulation of ROS and subsequently de-regulated mitosis and ultimately apoptosis in the NSPCs, leading to hydrocephalus development.
Figures
Similar articles
-
Oxidative stress-specific interaction between FANCD2 and FOXO3a.Blood. 2010 Feb 25;115(8):1545-8. doi: 10.1182/blood-2009-07-234385. Epub 2009 Dec 29. Blood. 2010. PMID: 20040763 Free PMC article.
-
Fancd2 is required for nuclear retention of Foxo3a in hematopoietic stem cell maintenance.J Biol Chem. 2015 Jan 30;290(5):2715-27. doi: 10.1074/jbc.M114.619536. Epub 2014 Dec 12. J Biol Chem. 2015. PMID: 25505262 Free PMC article.
-
Fanconi anemia links reactive oxygen species to insulin resistance and obesity.Antioxid Redox Signal. 2012 Oct 15;17(8):1083-98. doi: 10.1089/ars.2011.4417. Epub 2012 Jun 25. Antioxid Redox Signal. 2012. PMID: 22482891 Free PMC article.
-
Molecular pathogenesis of fanconi anemia.Int J Hematol. 2002 Feb;75(2):123-8. doi: 10.1007/BF02982016. Int J Hematol. 2002. PMID: 11939257 Review.
-
Fanconi anaemia proteins: major roles in cell protection against oxidative damage.Bioessays. 2003 Jun;25(6):589-95. doi: 10.1002/bies.10283. Bioessays. 2003. PMID: 12766948 Review.
Cited by
-
In Vivo RNAi Screen Unveils PPARγ as a Regulator of Hematopoietic Stem Cell Homeostasis.Stem Cell Reports. 2017 May 9;8(5):1242-1255. doi: 10.1016/j.stemcr.2017.03.008. Epub 2017 Apr 13. Stem Cell Reports. 2017. PMID: 28416286 Free PMC article.
-
Novel therapeutic modulators of astrocytes for hydrocephalus.Front Mol Neurosci. 2022 Sep 26;15:932955. doi: 10.3389/fnmol.2022.932955. eCollection 2022. Front Mol Neurosci. 2022. PMID: 36226316 Free PMC article. Review.
-
Edaravone reduces astrogliosis and apoptosis in young rats with kaolin-induced hydrocephalus.Childs Nerv Syst. 2017 Mar;33(3):419-428. doi: 10.1007/s00381-016-3313-x. Epub 2016 Dec 17. Childs Nerv Syst. 2017. PMID: 27988876
References
-
- Bani-Yaghoub M, Tremblay RG, Lei JX, Zhang D, Zurakowski B, Sandhu JK, Smith B, Ribecco-Lutkiewicz M, Kennedy J, Walker PR, and Sikorska M. Role of Sox2 in the development of the mouse neocortex. Dev Biol 295: 52–66, 2006 - PubMed
-
- Banizs B, Pike MM, Millican CL, et al. . Dysfunctional cilia lead to altered ependyma and choroid plexus function, and result in the formation of hydrocephalus. Development 132: 5329–5339, 2005 - PubMed
-
- Brunet A, Sweeney LB, Sturgill JF, et al. . Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science 303: 2011–2015, 2004 - PubMed
-
- Caner H, Atasever A, Kilinc K, Durgun B, Peker S, and Ozcan OE. Lipid peroxide level increase in experimental hydrocephalus. Acta Neurochir (Wien) 121: 68–71, 1993 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous