

Cultured primary human fetal retinal pigment epithelium (hfRPE) as a model for evaluating RPE metabolism
- PMID: 24485945
- PMCID: PMC4345411
- DOI: 10.1016/j.exer.2014.01.015
Cultured primary human fetal retinal pigment epithelium (hfRPE) as a model for evaluating RPE metabolism
Abstract
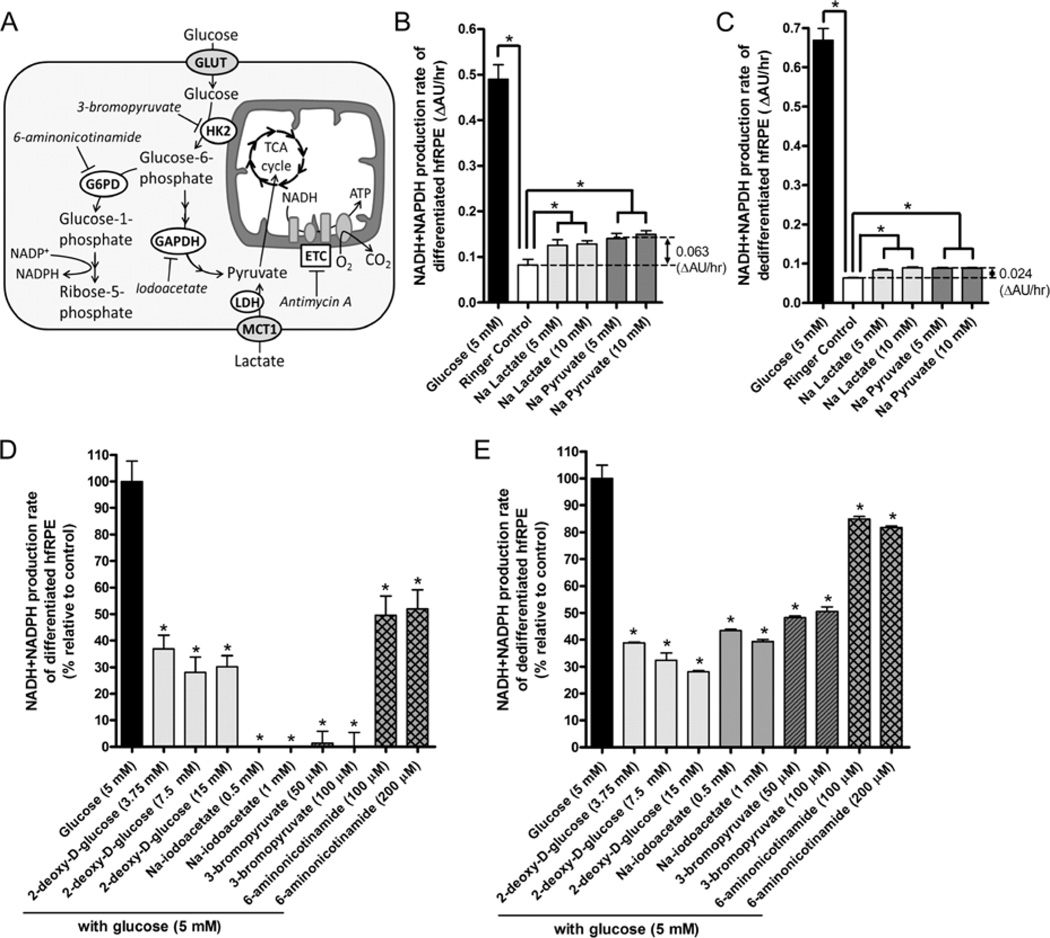
Mitochondrial dysfunction has been shown to contribute to age-related and proliferative retinal diseases. Over the past decade, the primary human fetal RPE (hfRPE) culture model has emerged as an effective tool for studying RPE function and mechanisms of retinal diseases. This model system has been rigorously characterized and shown to closely resemble native RPE cells at the genomic and protein level, and that they are capable of accomplishing the characteristic functions of a healthy native RPE (e.g., rod phagocytosis, ion and fluid transport, and retinoid cycle). In this review, we demonstrated that the metabolic activity of the RPE is an indicator of its health and state of differentiation, and present the hfRPE culture model as a valuable in vitro system for evaluating RPE metabolism in the context of RPE differentiation and retinal disease.
Keywords: differentiation; metabolism; primary cultured human fetal RPE.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures

References
-
- Adijanto J, Philp NJ. The SLC16A family of monocarboxylate transporters (MCTs)--physiology and function in cellular metabolism, pH homeostasis, and fluid transport. Current topics in membranes. 2012;70:275–311. - PubMed
-
- Agathocleous M, Harris WA. Metabolism in physiological cell proliferation and differentiation. Trends in cell biology. 2013;23:484–492. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
