

Tyr phosphorylation of PDP1 toggles recruitment between ACAT1 and SIRT3 to regulate the pyruvate dehydrogenase complex
- PMID: 24486017
- PMCID: PMC3943932
- DOI: 10.1016/j.molcel.2013.12.026
Tyr phosphorylation of PDP1 toggles recruitment between ACAT1 and SIRT3 to regulate the pyruvate dehydrogenase complex
Abstract
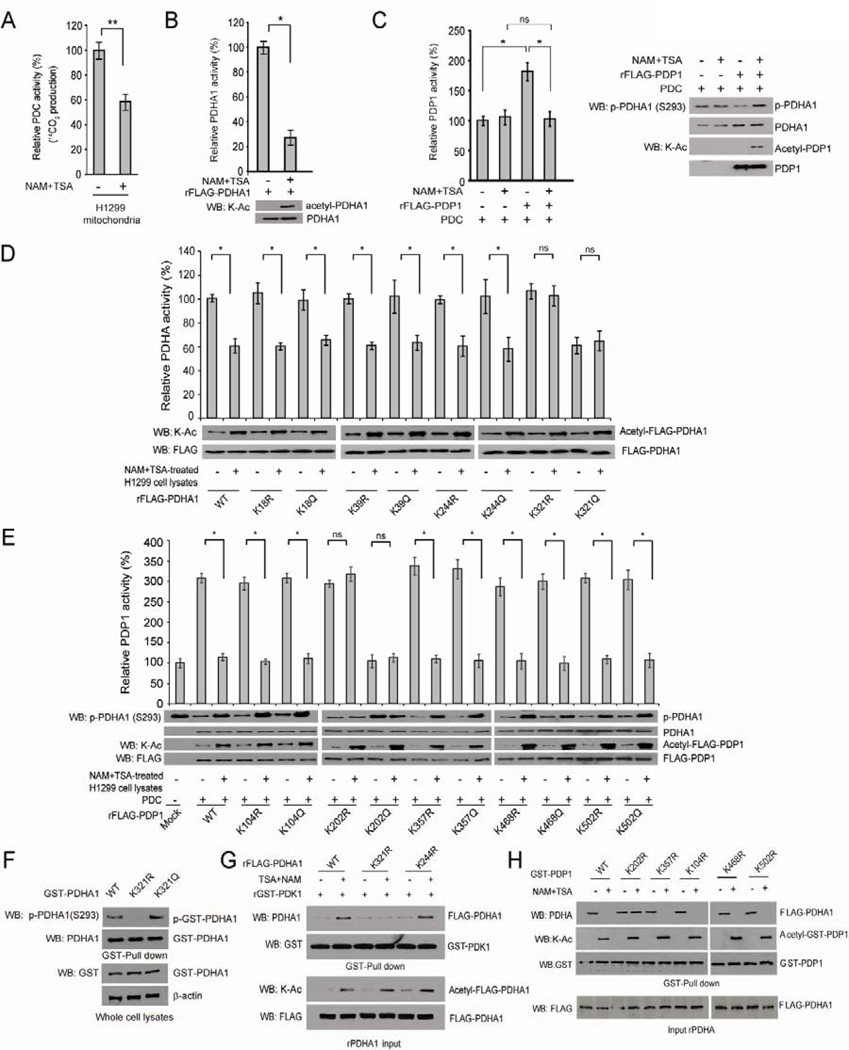
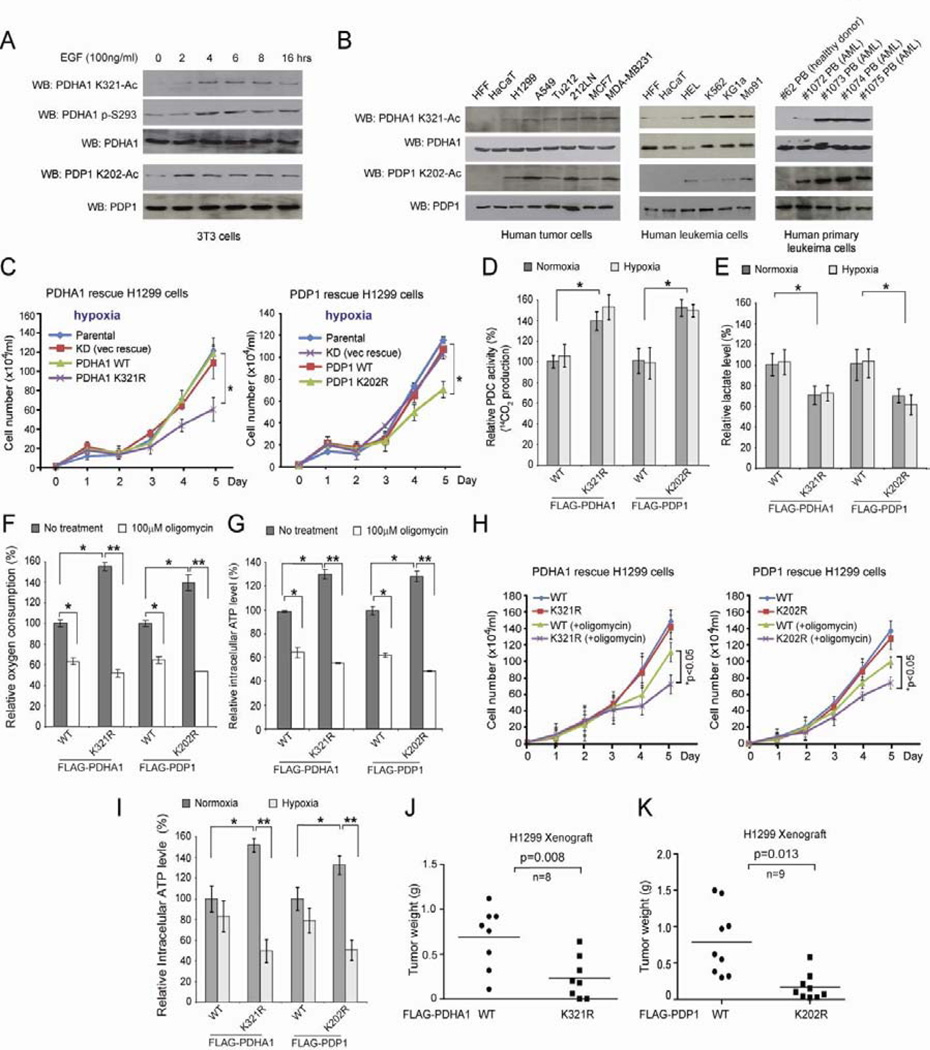
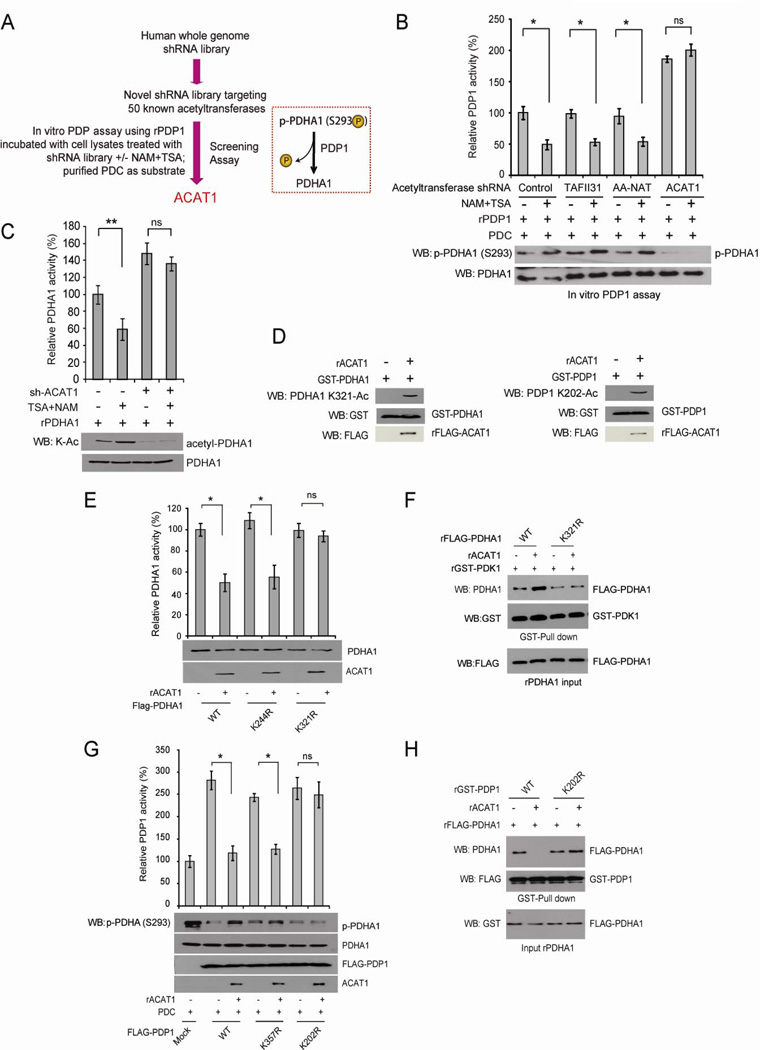
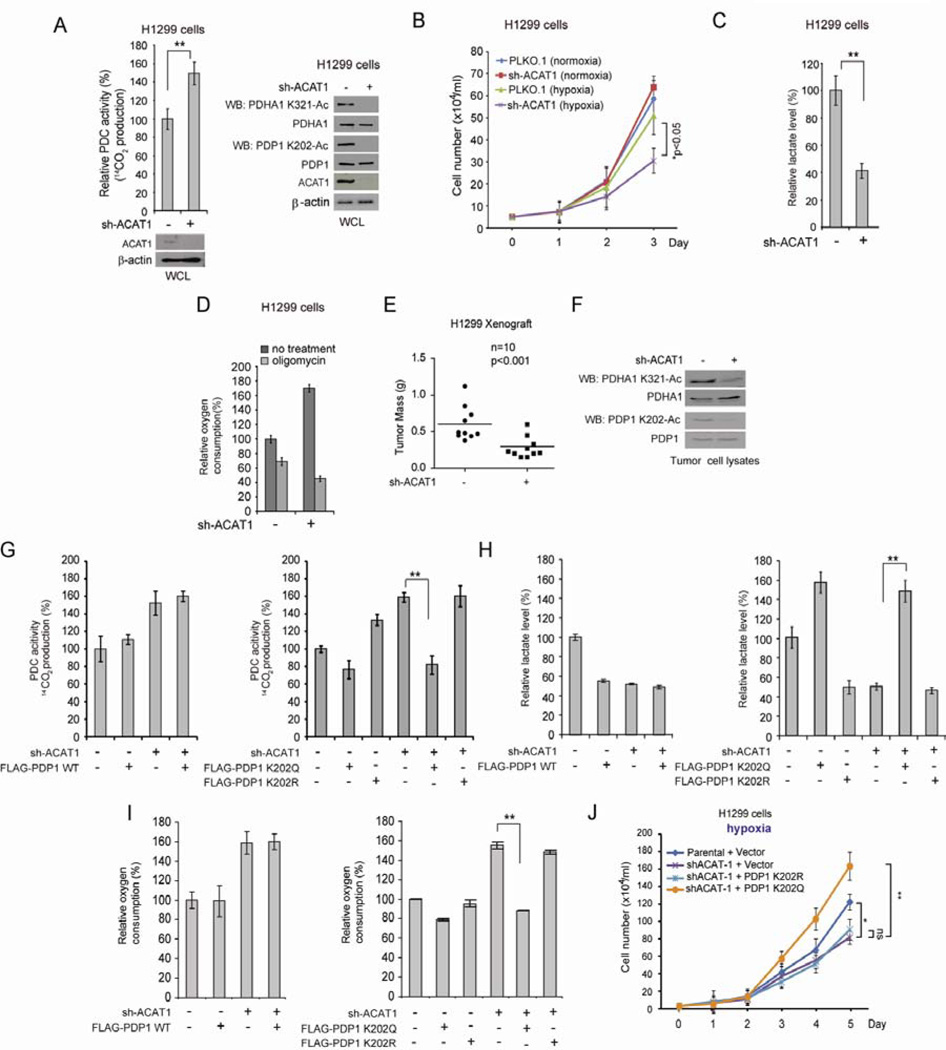
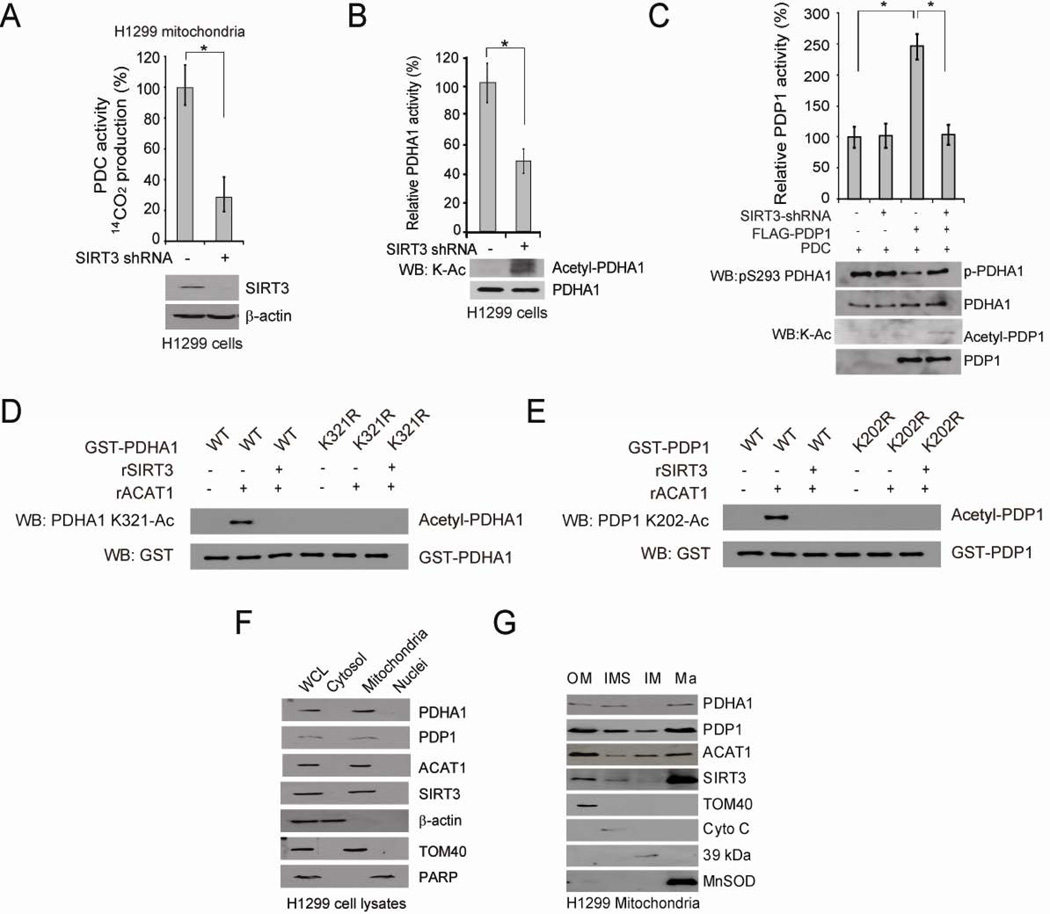
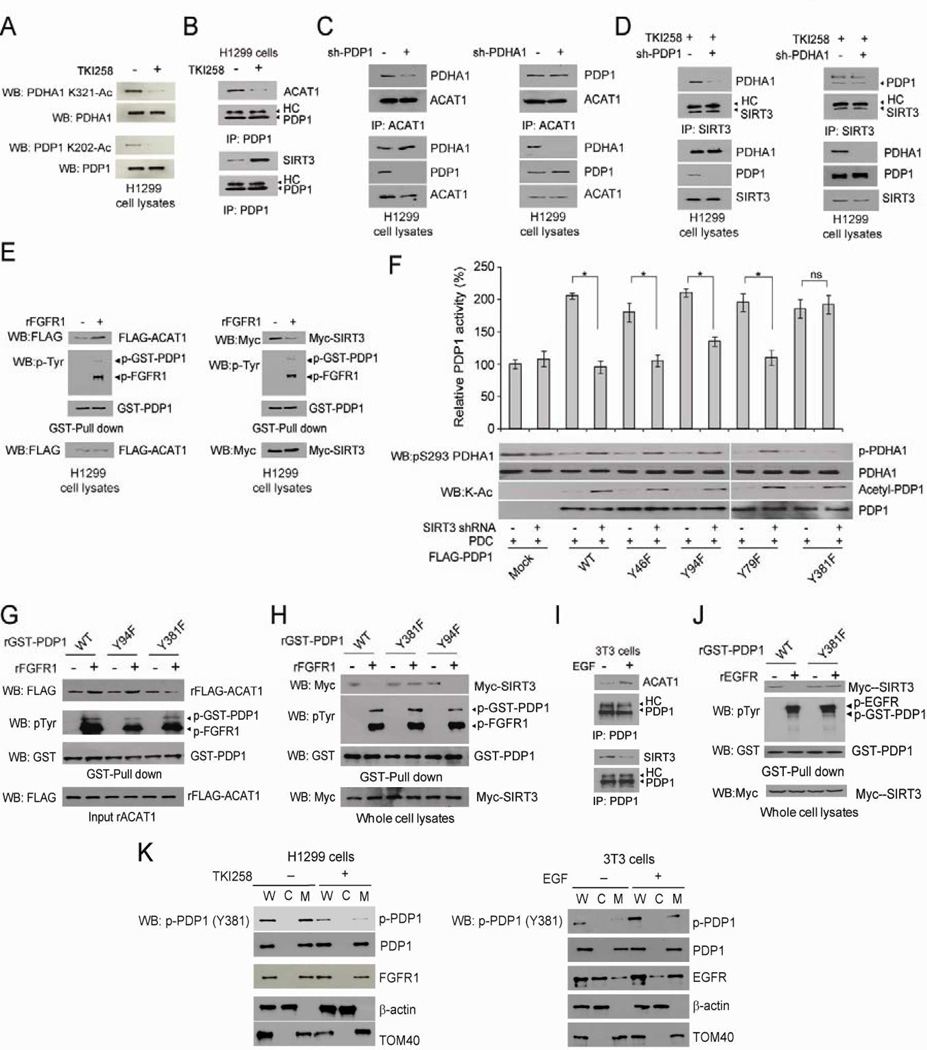
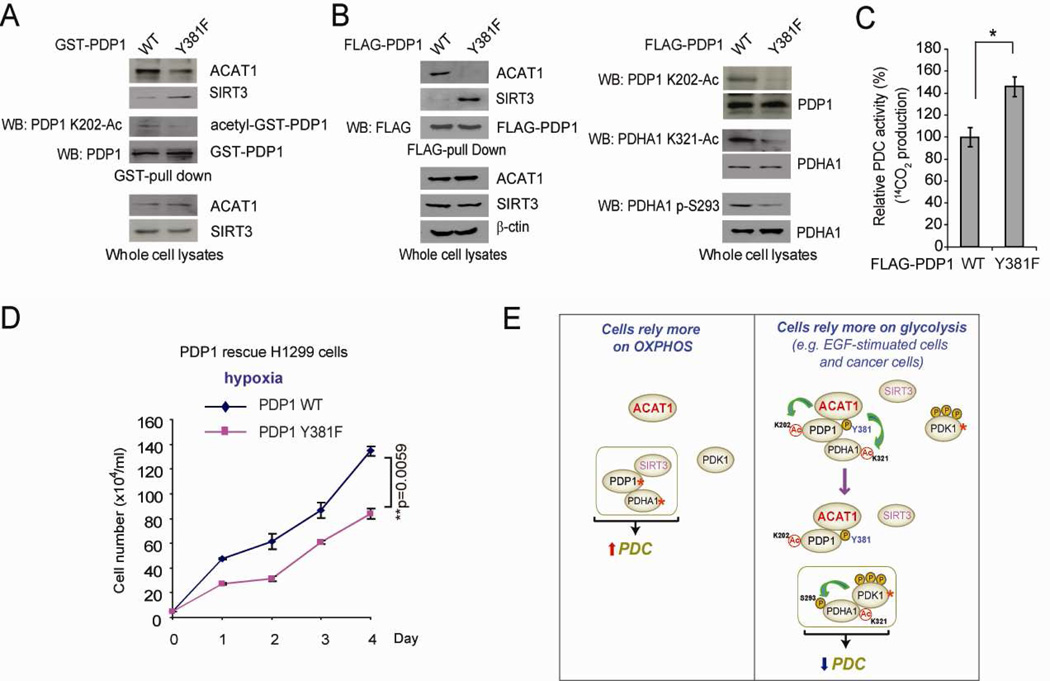
Mitochondrial pyruvate dehydrogenase complex (PDC) is crucial for glucose homeostasis in mammalian cells. The current understanding of PDC regulation involves inhibitory serine phosphorylation of pyruvate dehydrogenase (PDH) by PDH kinase (PDK), whereas dephosphorylation of PDH by PDH phosphatase (PDP) activates PDC. Here, we report that lysine acetylation of PDHA1 and PDP1 is common in epidermal growth factor (EGF)-stimulated cells and diverse human cancer cells. K321 acetylation inhibits PDHA1 by recruiting PDK1, and K202 acetylation inhibits PDP1 by dissociating its substrate PDHA1, both of which are important in promoting glycolysis in cancer cells and consequent tumor growth. Moreover, we identified mitochondrial ACAT1 and SIRT3 as the upstream acetyltransferase and deacetylase, respectively, of PDHA1 and PDP1, while knockdown of ACAT1 attenuates tumor growth. Furthermore, Y381 phosphorylation of PDP1 dissociates SIRT3 and recruits ACAT1 to PDC. Together, hierarchical, distinct posttranslational modifications act in concert to control molecular composition of PDC and contribute to the Warburg effect.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Balasse EO, Fery F. Ketone body production and disposal: effects of fasting, diabetes, and exercise. Diabetes Metab Rev. 1989;5:247–270. - PubMed
-
- Cairns RA, Harris IS, Mak TW. Regulation of cancer cell metabolism. Nature reviews. Cancer. 2011;11:85–95. - PubMed
-
- Choudhary C, Kumar C, Gnad F, Nielsen ML, Rehman M, Walther TC, Olsen JV, Mann M. Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science. 2009;325:834–840. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
