

Sustained and selective suppression of intestinal cholesterol synthesis by Ro 48-8071, an inhibitor of 2,3-oxidosqualene:lanosterol cyclase, in the BALB/c mouse
- PMID: 24486573
- PMCID: PMC3989900
- DOI: 10.1016/j.bcp.2014.01.031
Sustained and selective suppression of intestinal cholesterol synthesis by Ro 48-8071, an inhibitor of 2,3-oxidosqualene:lanosterol cyclase, in the BALB/c mouse
Abstract
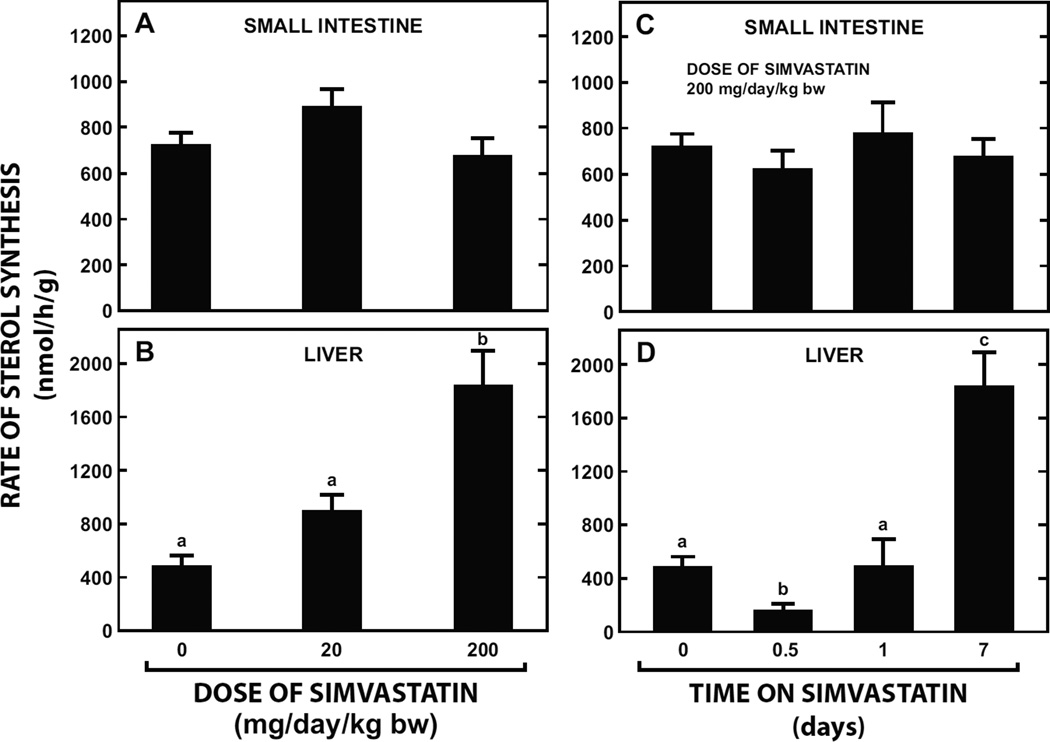
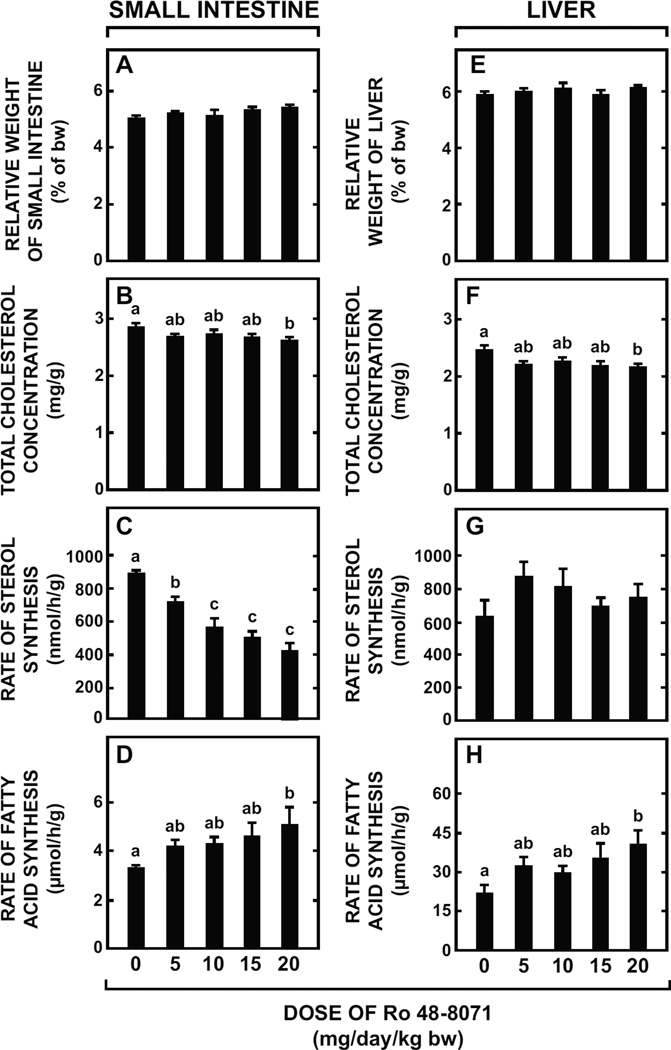
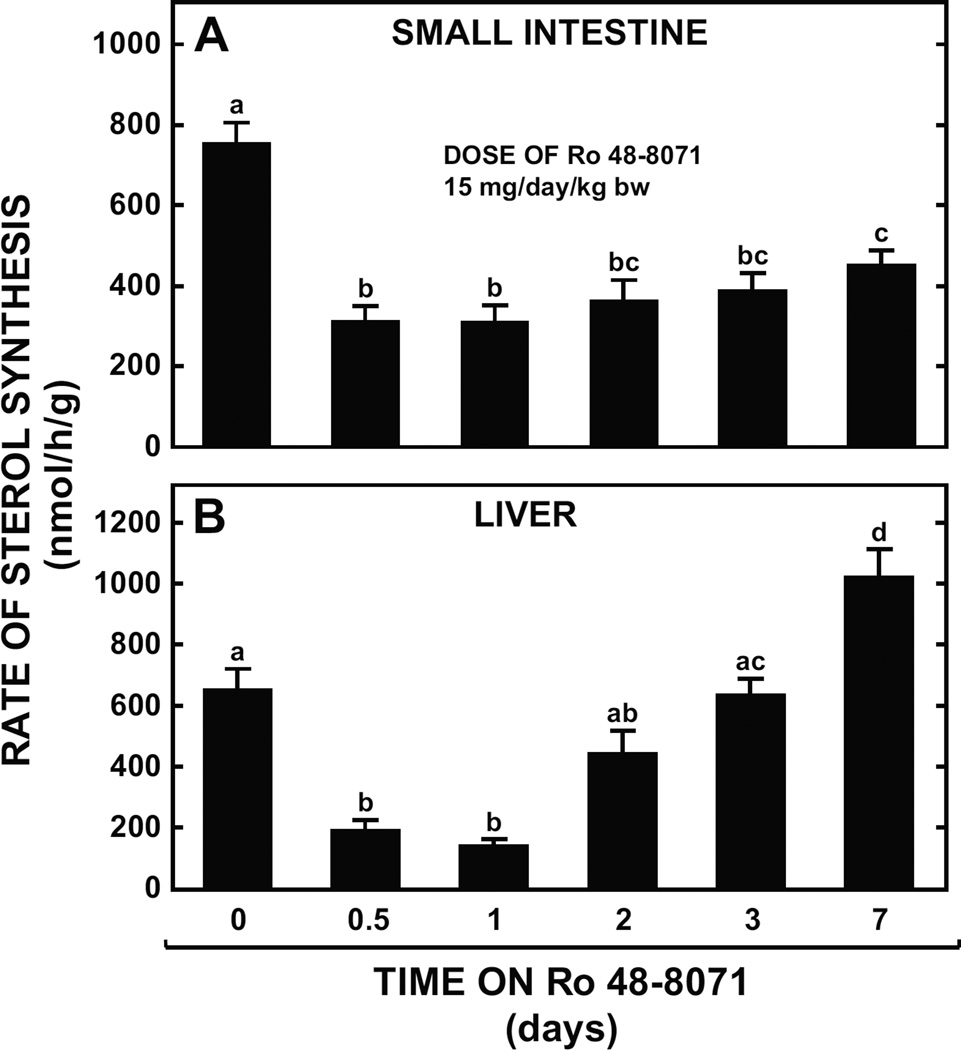
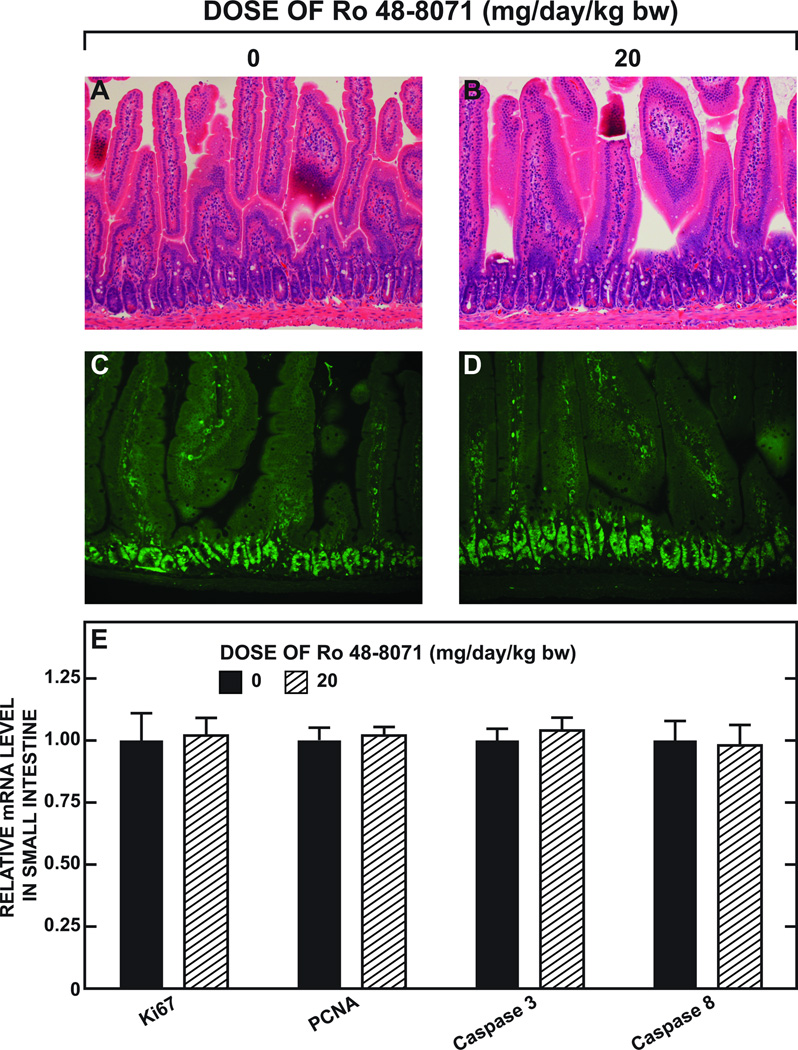
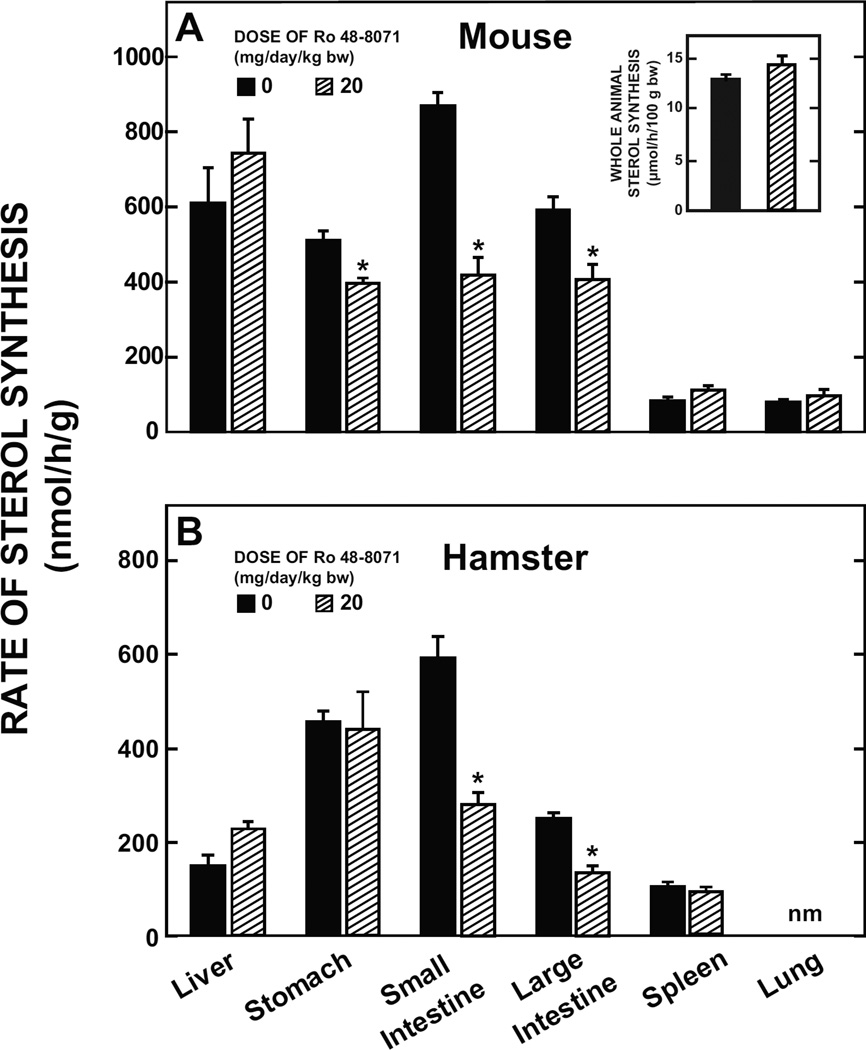
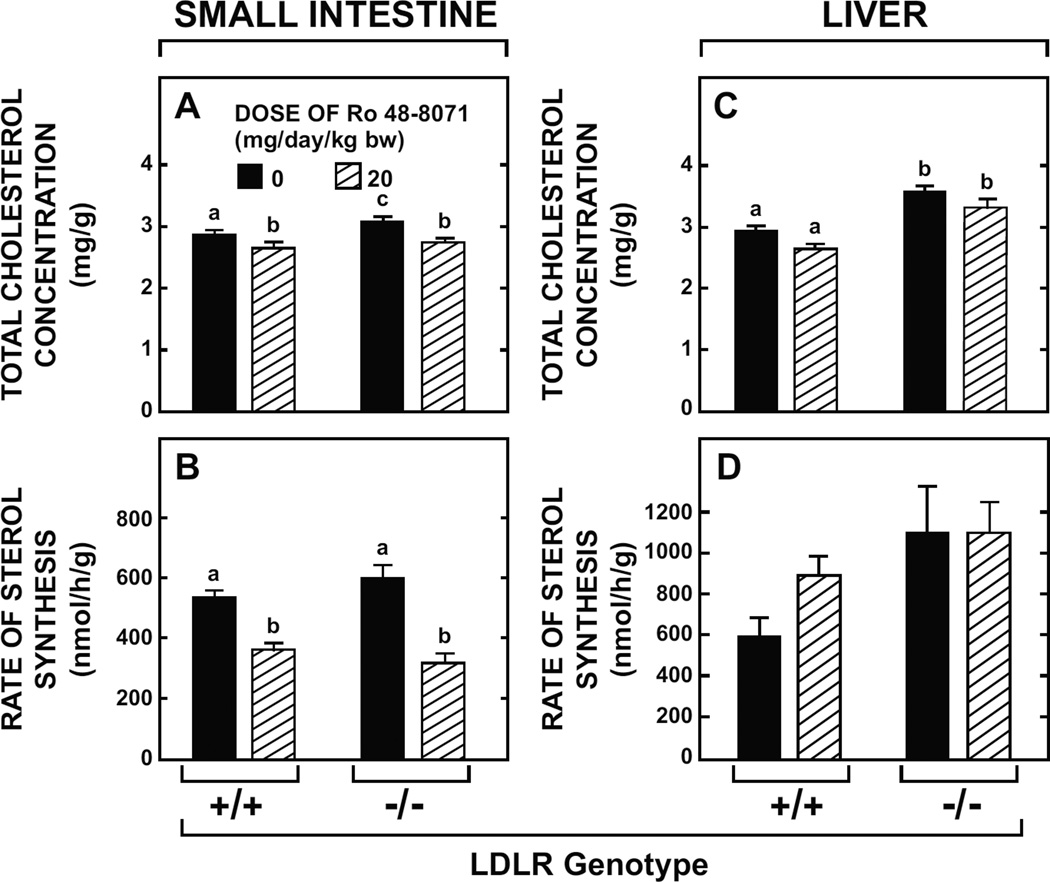
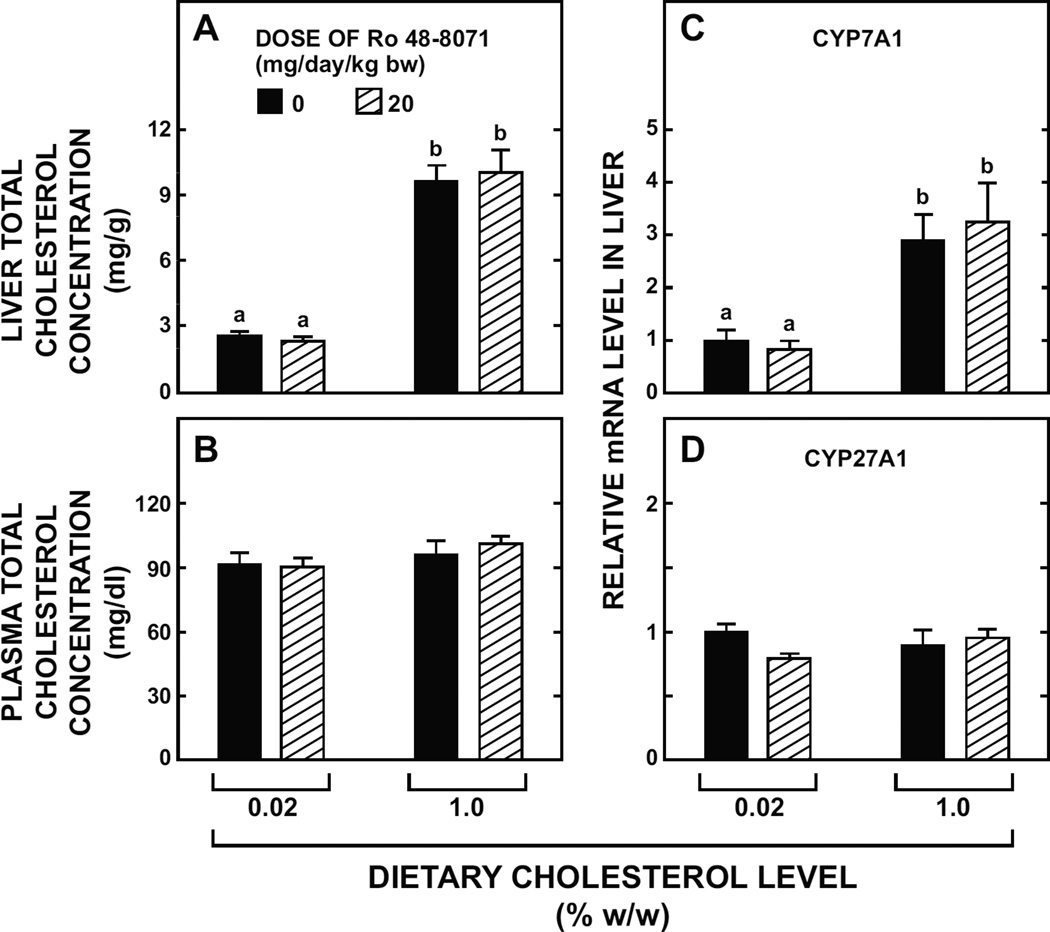
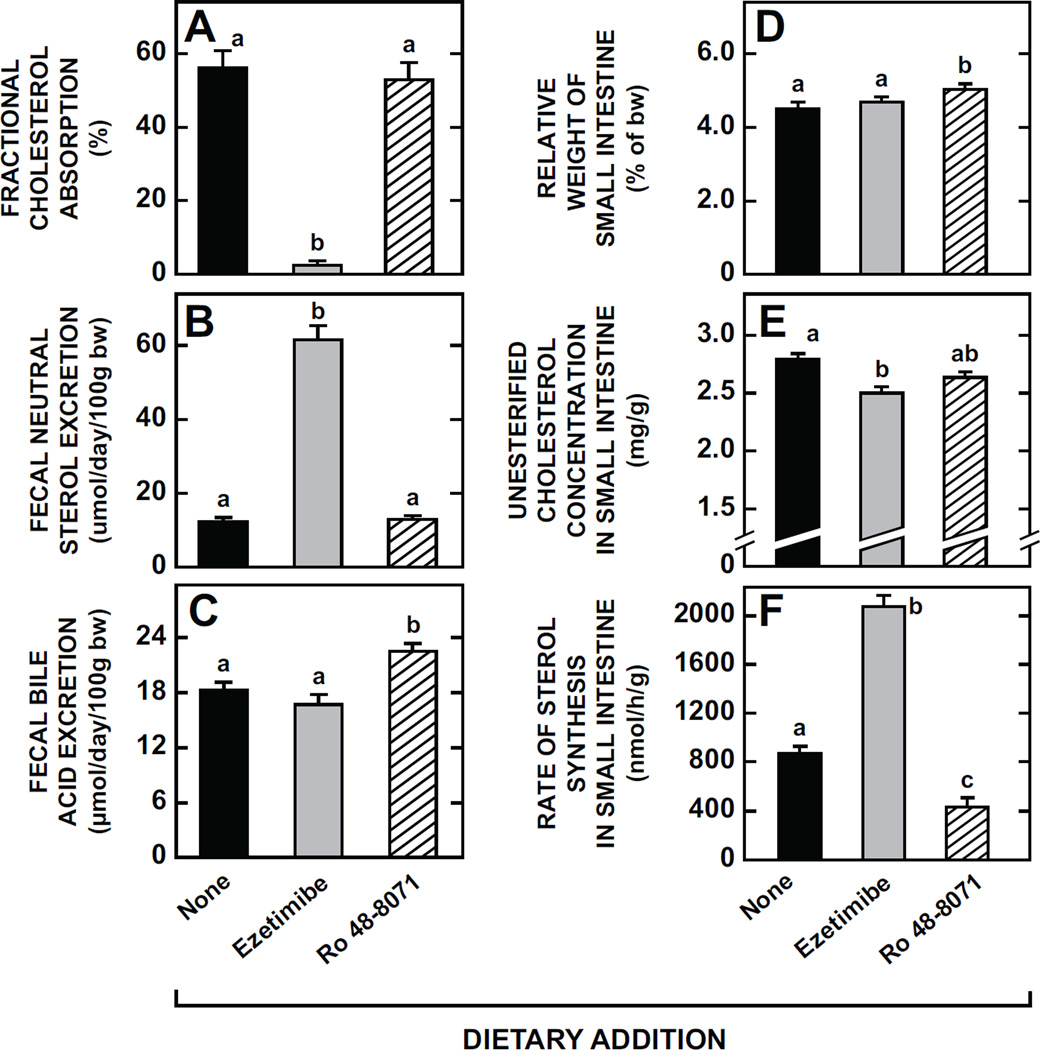
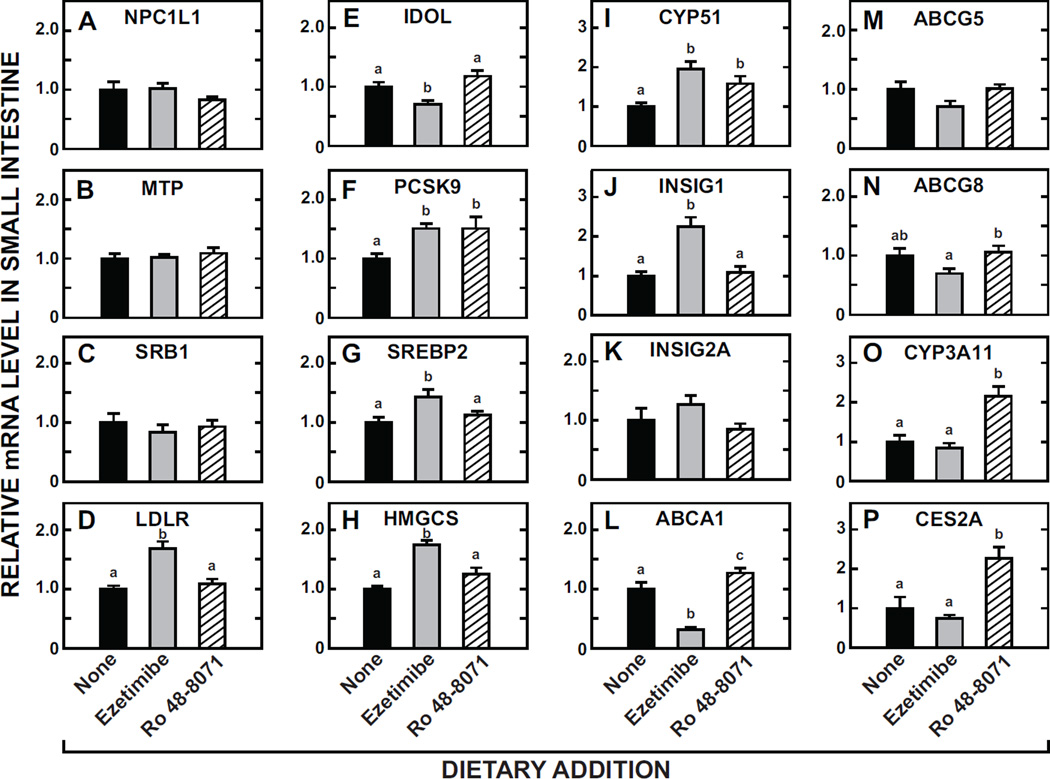
The small intestine plays a fundamentally important role in regulating whole body cholesterol balance and plasma lipoprotein composition. This is articulated through the interplay of a constellation of genes that ultimately determines the net amount of chylomicron cholesterol delivered to the liver. Major advances in our insights into regulation of the cholesterol absorption pathway have been made using genetically manipulated mouse models and agents such as ezetimibe. One unresolved question is how a sustained pharmacological inhibition of intestinal cholesterol synthesis in vivo may affect cholesterol handling by the absorptive cells. Here we show that the lanosterol cyclase inhibitor, Ro 48-8071, when fed to BALB/c mice in a chow diet (20 mg/day/kg body weight), leads to a rapid and sustained inhibition (>50%) of cholesterol synthesis in the whole small intestine. Sterol synthesis was also reduced in the large intestine and stomach. In contrast, hepatic cholesterol synthesis, while markedly suppressed initially, rebounded to higher than baseline rates within 7 days. Whole body cholesterol synthesis, fractional cholesterol absorption, and fecal neutral and acidic sterol excretion were not consistently changed with Ro 48-8071 treatment. There were no discernible effects of this agent on intestinal histology as determined by H&E staining and the level of Ki67, an index of proliferation. The mRNA expression for multiple genes involved in intestinal cholesterol regulation including NPC1L1 was mostly unchanged although there was a marked rise in the mRNA level for the PXR target genes CYP3A11 and CES2A.
Keywords: Cholesterol absorption; Enterocyte; Ezetimibe; Liver; Statins.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Grundy SM. Absorption and metabolism of dietary cholesterol. Annu Rev Nutr. 1983;3:71–96. - PubMed
-
- Stange EF, Dietschy JM. Cholesterol Absorption and Metabolism by the Intestinal Epithelium. In: Danielsson H, Sjövall J, editors. Sterols and Bile Acids. New York: Elsevier Science Publishers; 1985. pp. 121–149.
-
- Turley SD, Dietschy JM. The Metabolism and Excretion of Cholesterol by the Liver. In: Arias IM, Jakoby WB, Popper H, Schachter D, Shafritz DA, editors. The Liver: Biology and Pathobiology. New York: Raven Press; 1988. pp. 617–641.
-
- Cooper AD. Hepatic uptake of chylomicron remnants. J Lipid Res. 1997;38:2173–2192. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
