

Cortical activity in the null space: permitting preparation without movement
- PMID: 24487233
- PMCID: PMC3955357
- DOI: 10.1038/nn.3643
Cortical activity in the null space: permitting preparation without movement
Abstract
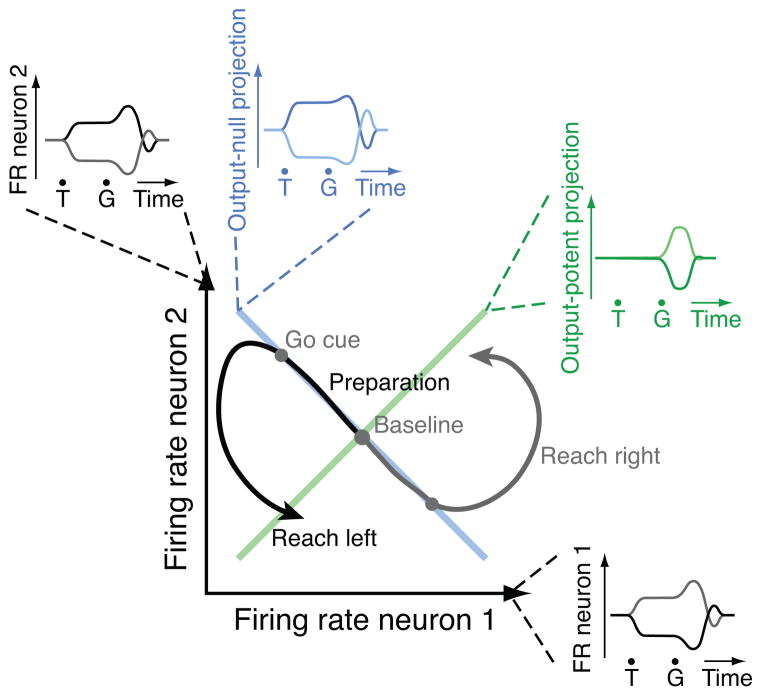
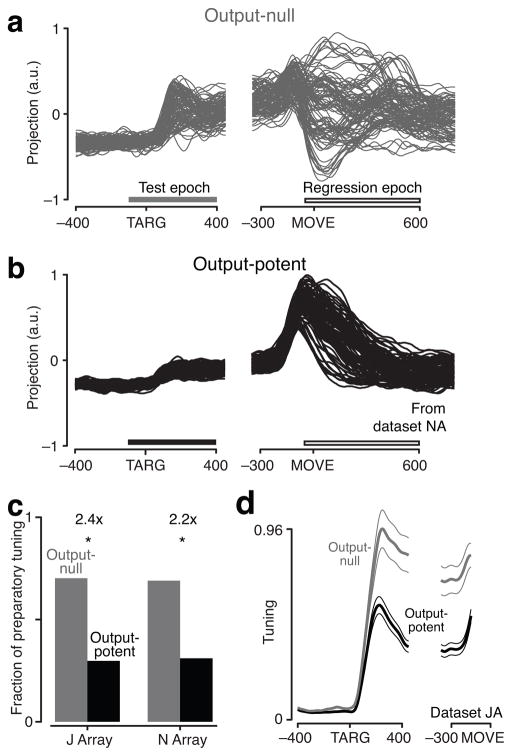
Neural circuits must perform computations and then selectively output the results to other circuits. Yet synapses do not change radically at millisecond timescales. A key question then is: how is communication between neural circuits controlled? In motor control, brain areas directly involved in driving movement are active well before movement begins. Muscle activity is some readout of neural activity, yet it remains largely unchanged during preparation. Here we find that during preparation, while the monkey holds still, changes in motor cortical activity cancel out at the level of these population readouts. Motor cortex can thereby prepare the movement without prematurely causing it. Further, we found evidence that this mechanism also operates in dorsal premotor cortex, largely accounting for how preparatory activity is attenuated in primary motor cortex. Selective use of 'output-null' vs. 'output-potent' patterns of activity may thus help control communication to the muscles and between these brain areas.
Figures





Comment in
-
Crouching tiger, hidden dimensions.Nat Neurosci. 2014 Mar;17(3):338-40. doi: 10.1038/nn.3663. Nat Neurosci. 2014. PMID: 24569828 No abstract available.
-
Commentary: Cortical activity in the null space: permitting preparation without movement.Front Neurosci. 2017 Sep 13;11:502. doi: 10.3389/fnins.2017.00502. eCollection 2017. Front Neurosci. 2017. PMID: 29503605 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
