

Mutant p53-associated myosin-X upregulation promotes breast cancer invasion and metastasis
- PMID: 24487586
- PMCID: PMC3934176
- DOI: 10.1172/JCI67280
Mutant p53-associated myosin-X upregulation promotes breast cancer invasion and metastasis
Erratum in
-
Corrigendum to Mutant p53-associated myosin-X upregulation promotes breast cancer invasion and metastasis.J Clin Invest. 2025 Dec 1;135(23):e201379. doi: 10.1172/JCI201379. eCollection 2025 Dec 1. J Clin Invest. 2025. PMID: 41321321 Free PMC article. No abstract available.
Abstract
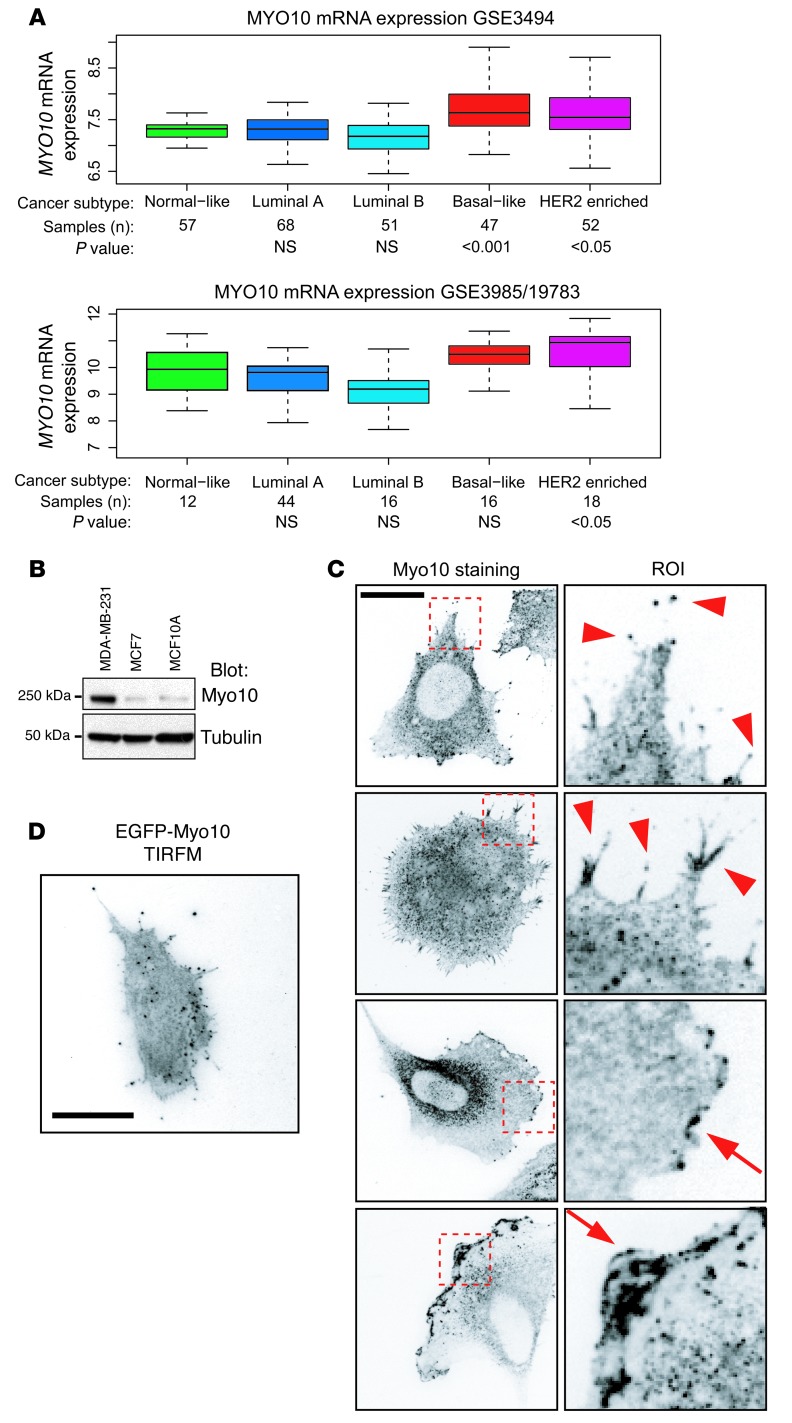
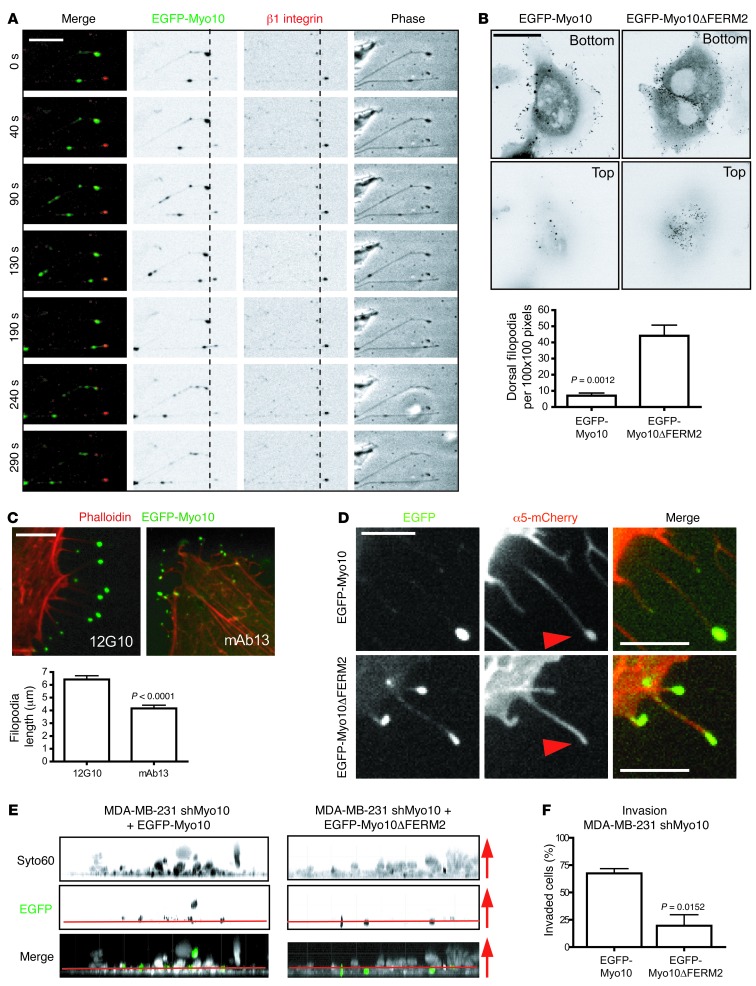
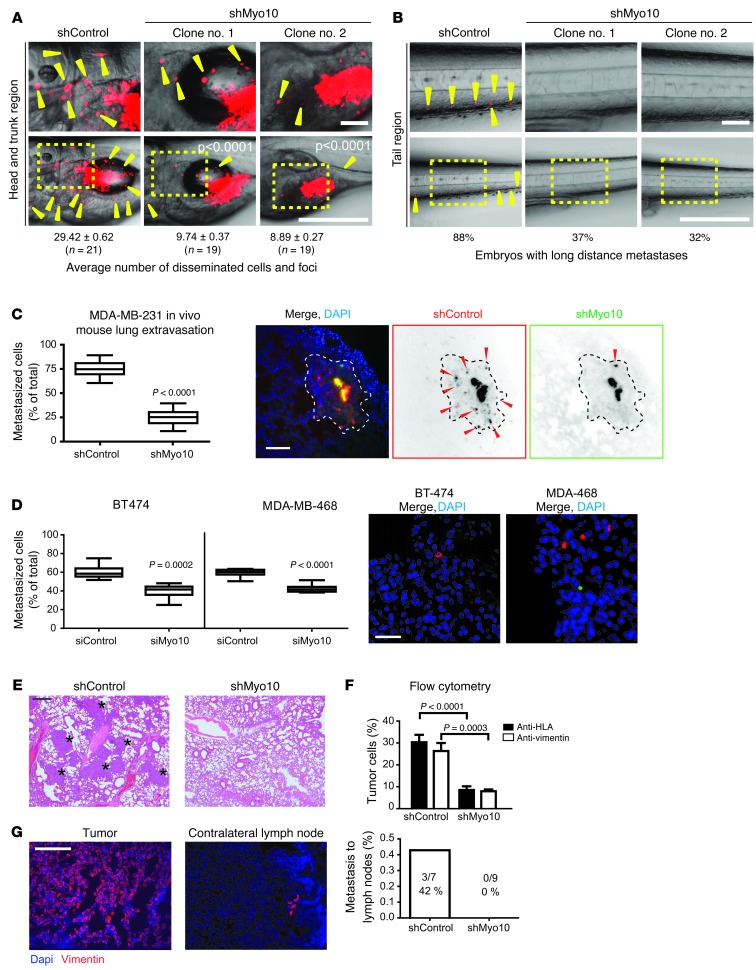
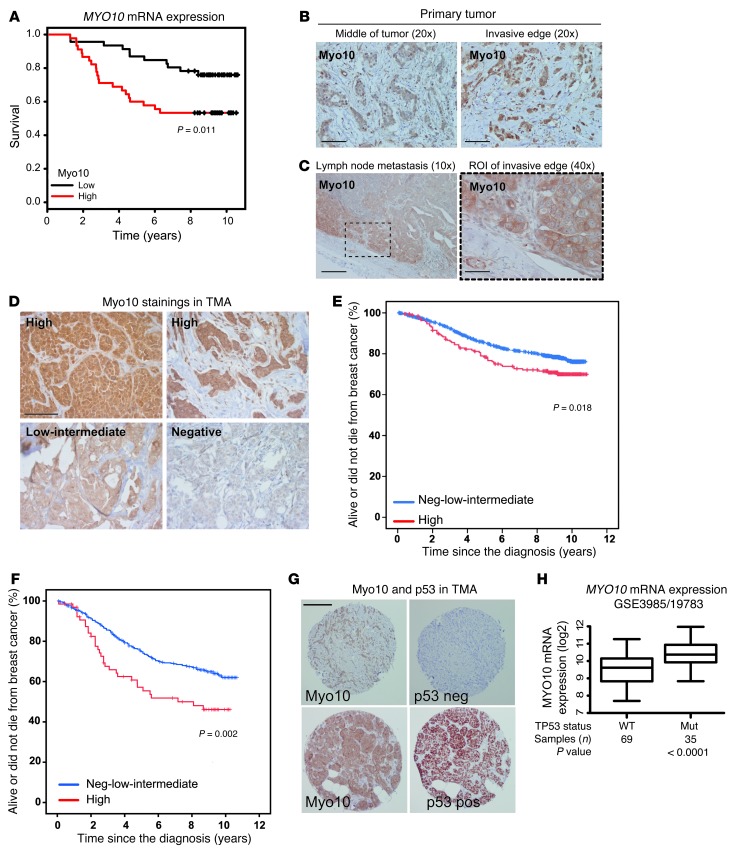
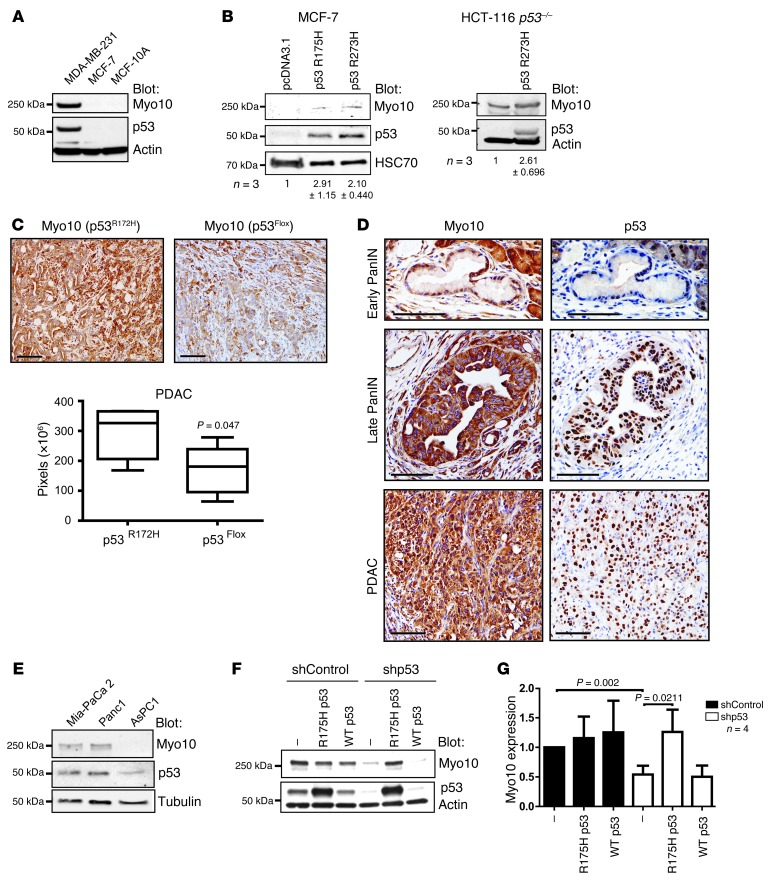
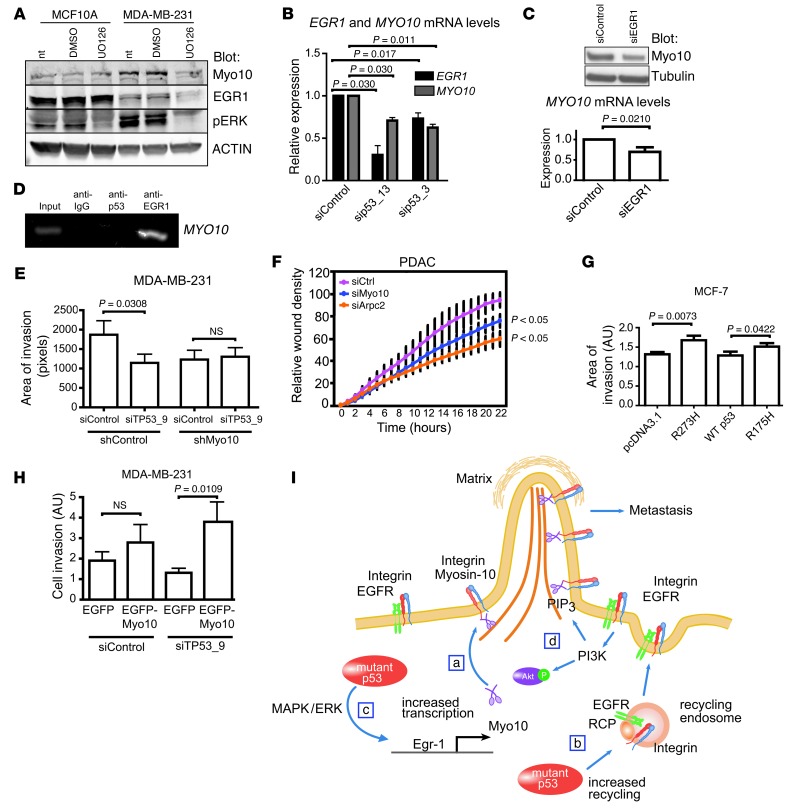
Mutations of the tumor suppressor TP53 are present in many forms of human cancer and are associated with increased tumor cell invasion and metastasis. Several mechanisms have been identified for promoting dissemination of cancer cells with TP53 mutations, including increased targeting of integrins to the plasma membrane. Here, we demonstrate a role for the filopodia-inducing motor protein Myosin-X (Myo10) in mutant p53-driven cancer invasion. Analysis of gene expression profiles from 2 breast cancer data sets revealed that MYO10 was highly expressed in aggressive cancer subtypes. Myo10 was required for breast cancer cell invasion and dissemination in multiple cancer cell lines and murine models of cancer metastasis. Evaluation of a Myo10 mutant without the integrin-binding domain revealed that the ability of Myo10 to transport β₁ integrins to the filopodia tip is required for invasion. Introduction of mutant p53 promoted Myo10 expression in cancer cells and pancreatic ductal adenocarcinoma in mice, whereas suppression of endogenous mutant p53 attenuated Myo10 levels and cell invasion. In clinical breast carcinomas, Myo10 was predominantly expressed at the invasive edges and correlated with the presence of TP53 mutations and poor prognosis. These data indicate that Myo10 upregulation in mutant p53-driven cancers is necessary for invasion and that plasma-membrane protrusions, such as filopodia, may serve as specialized metastatic engines.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
