

The coordination of C4 photosynthesis and the CO2-concentrating mechanism in maize and Miscanthus x giganteus in response to transient changes in light quality
- PMID: 24488966
- PMCID: PMC3938620
- DOI: 10.1104/pp.113.224683
The coordination of C4 photosynthesis and the CO2-concentrating mechanism in maize and Miscanthus x giganteus in response to transient changes in light quality
Abstract
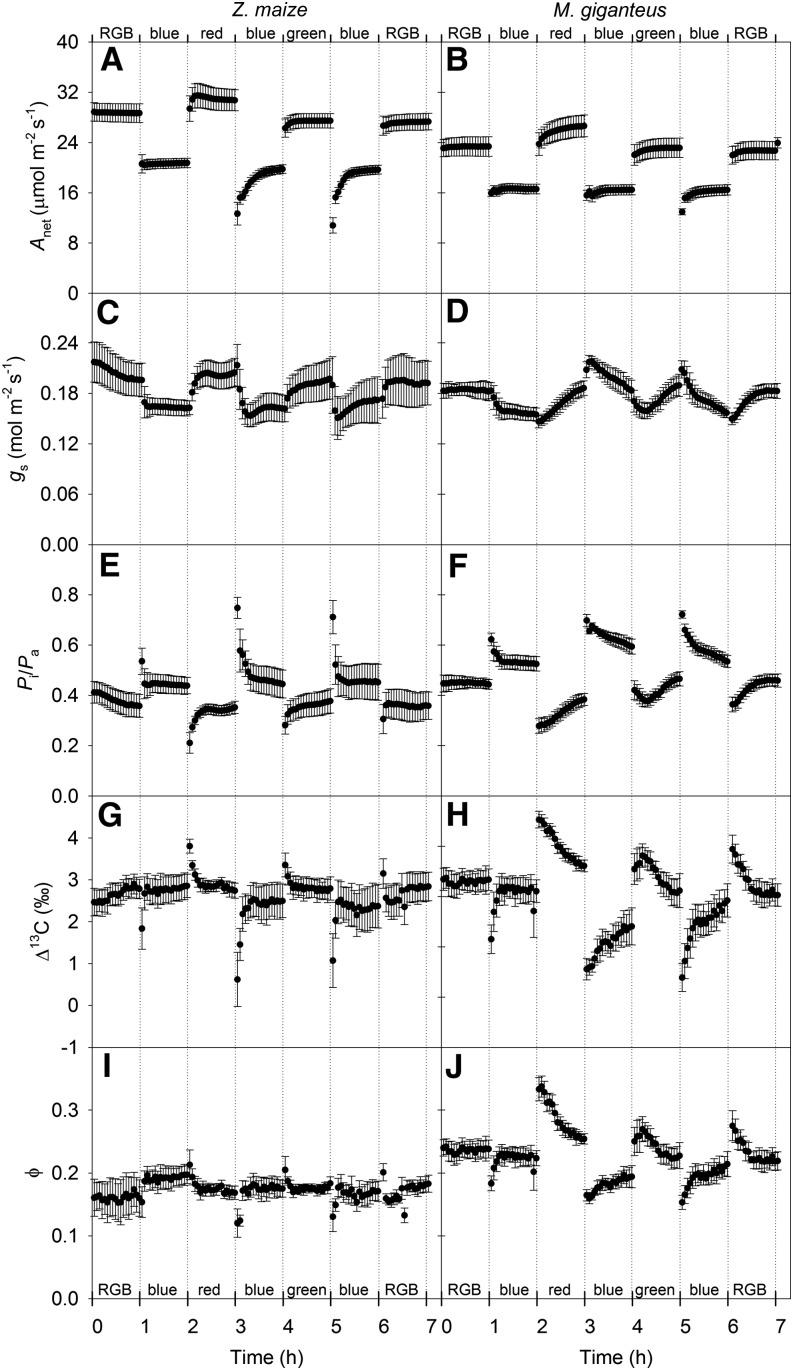
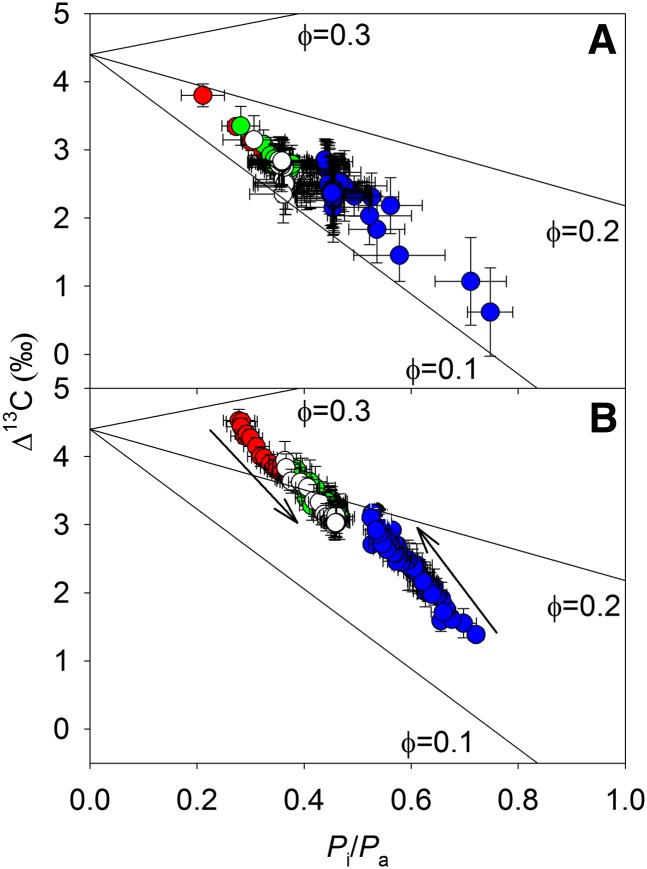
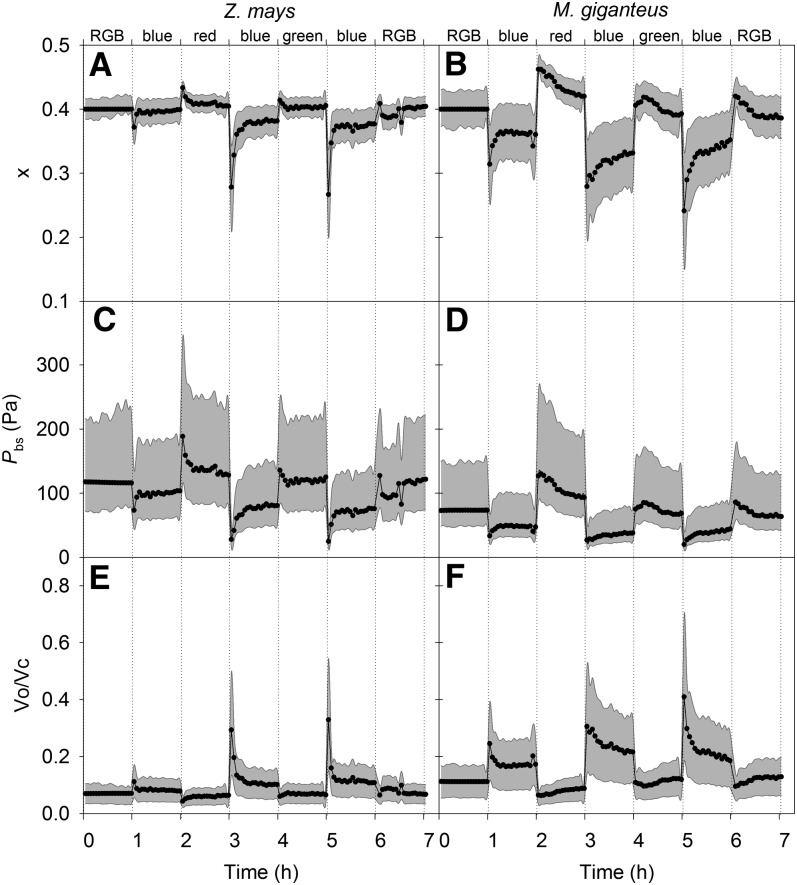
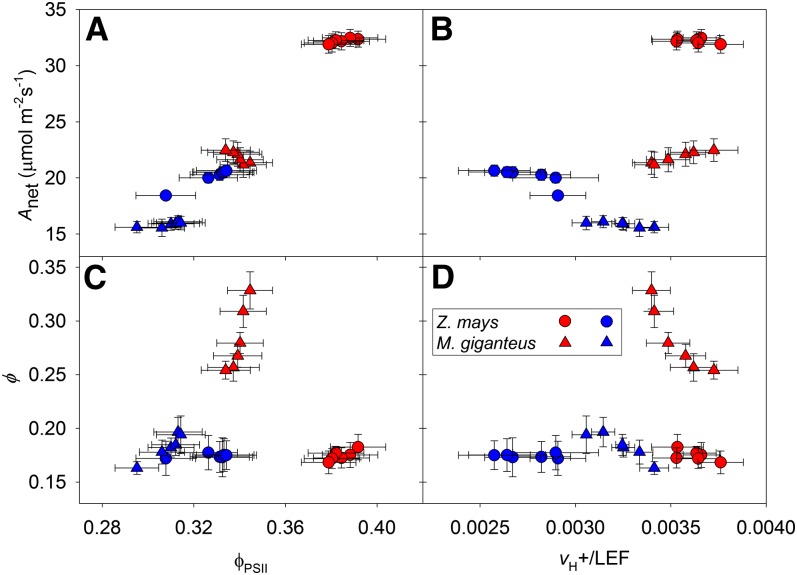
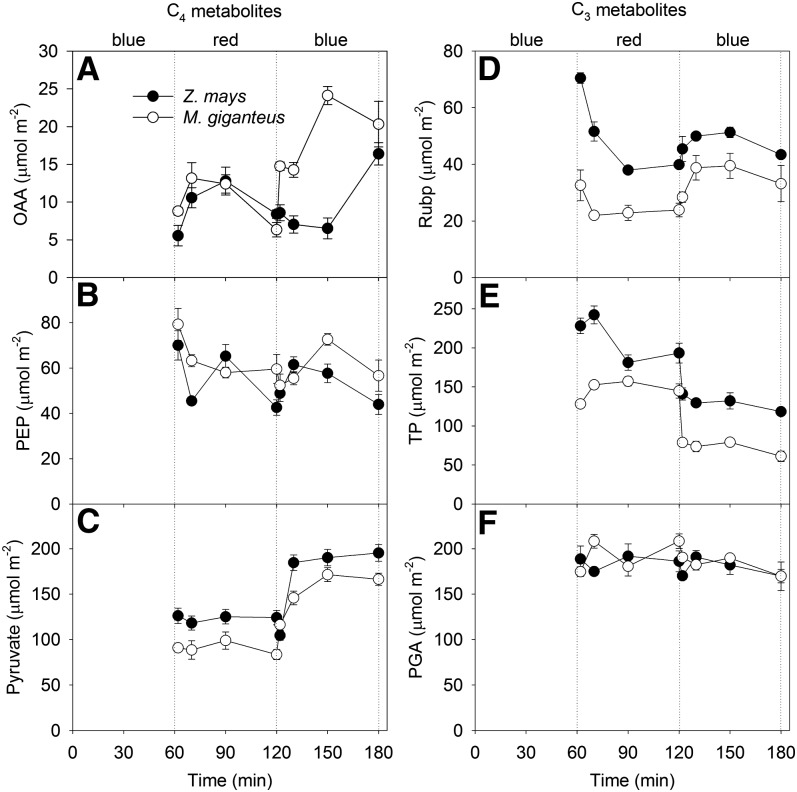
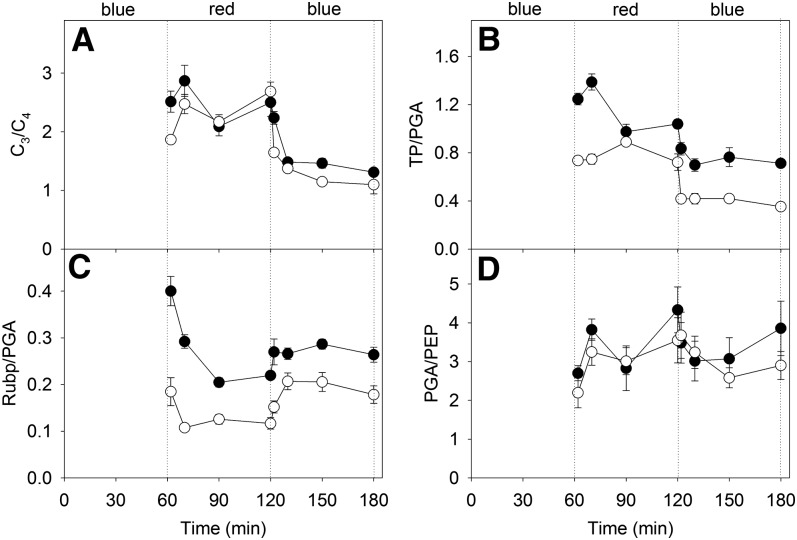
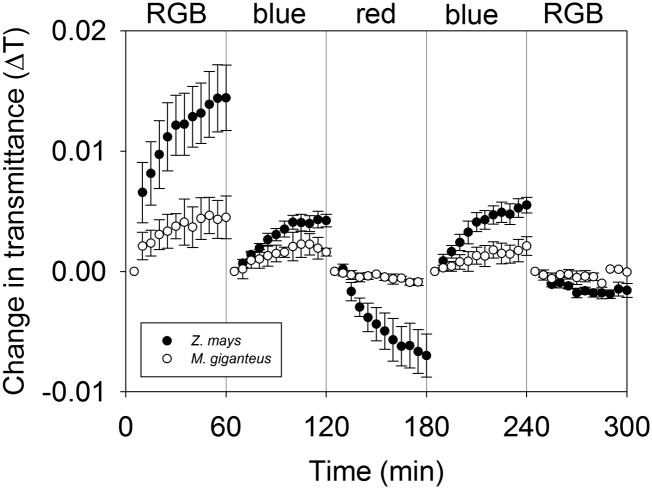
Unequal absorption of photons between photosystems I and II, and between bundle-sheath and mesophyll cells, are likely to affect the efficiency of the CO2-concentrating mechanism in C4 plants. Under steady-state conditions, it is expected that the biochemical distribution of energy (ATP and NADPH) and photosynthetic metabolite concentrations will adjust to maintain the efficiency of C4 photosynthesis through the coordination of the C3 (Calvin-Benson-Bassham) and C4 (CO2 pump) cycles. However, under transient conditions, changes in light quality will likely alter the coordination of the C3 and C4 cycles, influencing rates of CO2 assimilation and decreasing the efficiency of the CO2-concentrating mechanism. To test these hypotheses, we measured leaf gas exchange, leaf discrimination, chlorophyll fluorescence, electrochromatic shift, photosynthetic metabolite pools, and chloroplast movement in maize (Zea mays) and Miscanthus × giganteus following transitional changes in light quality. In both species, the rate of net CO2 assimilation responded quickly to changes in light treatments, with lower rates of net CO2 assimilation under blue light compared with red, green, and blue light, red light, and green light. Under steady state, the efficiency of CO2-concentrating mechanisms was similar; however, transient changes affected the coordination of C3 and C4 cycles in M. giganteus but to a lesser extent in maize. The species differences in the ability to coordinate the activities of C3 and C4 cycles appear to be related to differences in the response of cyclic electron flux around photosystem I and potentially chloroplast rearrangement in response to changes in light quality.
Figures
Similar articles
-
The influence of light quality on C4 photosynthesis under steady-state conditions in Zea mays and Miscanthus×giganteus: changes in rates of photosynthesis but not the efficiency of the CO2 concentrating mechanism.Plant Cell Environ. 2012 May;35(5):982-93. doi: 10.1111/j.1365-3040.2011.02466.x. Epub 2011 Dec 13. Plant Cell Environ. 2012. PMID: 22082455
-
Photosynthetic flexibility in maize exposed to salinity and shade.J Exp Bot. 2014 Jul;65(13):3715-24. doi: 10.1093/jxb/eru130. Epub 2014 Apr 1. J Exp Bot. 2014. PMID: 24692650 Free PMC article.
-
The efficiency of C4 photosynthesis under low light conditions in Zea mays, Miscanthus x giganteus and Flaveria bidentis.Plant Cell Environ. 2013 Feb;36(2):365-81. doi: 10.1111/j.1365-3040.2012.02579.x. Epub 2012 Aug 15. Plant Cell Environ. 2013. PMID: 22812384
-
Bundle-sheath leakiness in C4 photosynthesis: a careful balancing act between CO2 concentration and assimilation.J Exp Bot. 2014 Jul;65(13):3443-57. doi: 10.1093/jxb/eru157. Epub 2014 Apr 22. J Exp Bot. 2014. PMID: 24755278 Review.
-
Prospects for improving CO2 fixation in C3-crops through understanding C4-Rubisco biogenesis and catalytic diversity.Curr Opin Plant Biol. 2016 Jun;31:135-42. doi: 10.1016/j.pbi.2016.04.002. Epub 2016 Apr 27. Curr Opin Plant Biol. 2016. PMID: 27131319 Review.
Cited by
-
The Coordination of Gene Expression within Photosynthesis Pathway for Acclimation of C4 Energy Crop Miscanthus lutarioriparius.Front Plant Sci. 2016 Feb 9;7:109. doi: 10.3389/fpls.2016.00109. eCollection 2016. Front Plant Sci. 2016. PMID: 26904072 Free PMC article.
-
Subdivision of Light Signaling Networks Contributes to Partitioning of C4 Photosynthesis.Plant Physiol. 2020 Mar;182(3):1297-1309. doi: 10.1104/pp.19.01053. Epub 2019 Dec 20. Plant Physiol. 2020. PMID: 31862840 Free PMC article.
-
The role of chloroplast movement in C4 photosynthesis: a theoretical analysis using a three-dimensional reaction-diffusion model for maize.J Exp Bot. 2023 Aug 3;74(14):4125-4142. doi: 10.1093/jxb/erad138. J Exp Bot. 2023. PMID: 37083863 Free PMC article.
-
Transgenic maize phosphoenolpyruvate carboxylase alters leaf-atmosphere CO2 and 13CO2 exchanges in Oryza sativa.Photosynth Res. 2019 Nov;142(2):153-167. doi: 10.1007/s11120-019-00655-4. Epub 2019 Jul 19. Photosynth Res. 2019. PMID: 31325077 Free PMC article.
-
Effects of elevated CO2 on competition between native and invasive grasses.Oecologia. 2020 Apr;192(4):1099-1110. doi: 10.1007/s00442-020-04636-6. Epub 2020 Apr 6. Oecologia. 2020. PMID: 32253494
References
-
- Badger MR, Sharkey TD, von Caemmerer S. (1984) The relationship between steady-state gas exchange of bean leaves and the levels of carbon-reduction-cycle intermediates. Planta 160: 305–313 - PubMed
-
- Bowling DR, Sargent SD, Tanner BD, Ehleringer JR. (2003) Tunable diode laser absorption spectroscopy for stable isotope studies of ecosystem-atmosphere CO2 exchange. Agric For Meteorol 118: 1–19
-
- Brown WV, Smith BN. (1972) Grass evolution, the Kranz syndrome, 13C/12C ratios, and continental drift. Nature 239: 345–346 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
