

Elovl2 ablation demonstrates that systemic DHA is endogenously produced and is essential for lipid homeostasis in mice
- PMID: 24489111
- PMCID: PMC3966705
- DOI: 10.1194/jlr.M046151
Elovl2 ablation demonstrates that systemic DHA is endogenously produced and is essential for lipid homeostasis in mice
Abstract
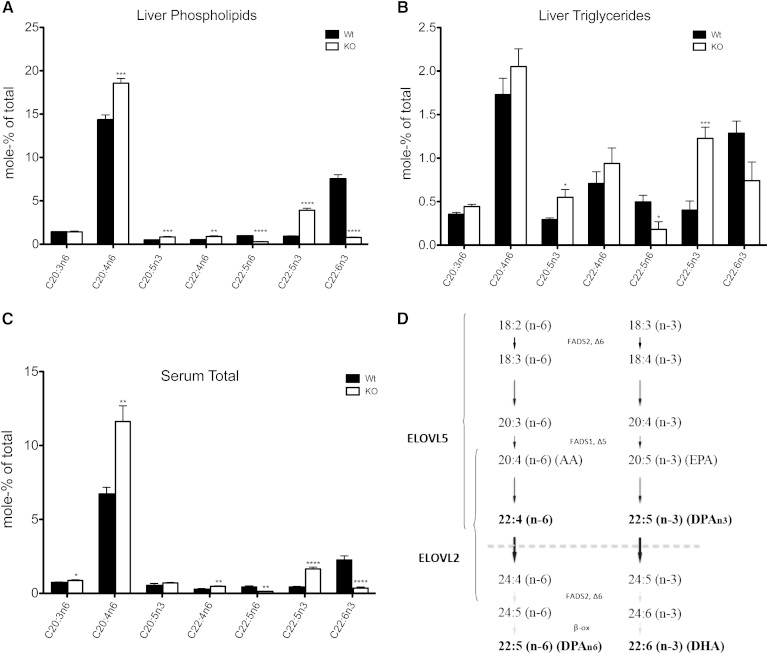
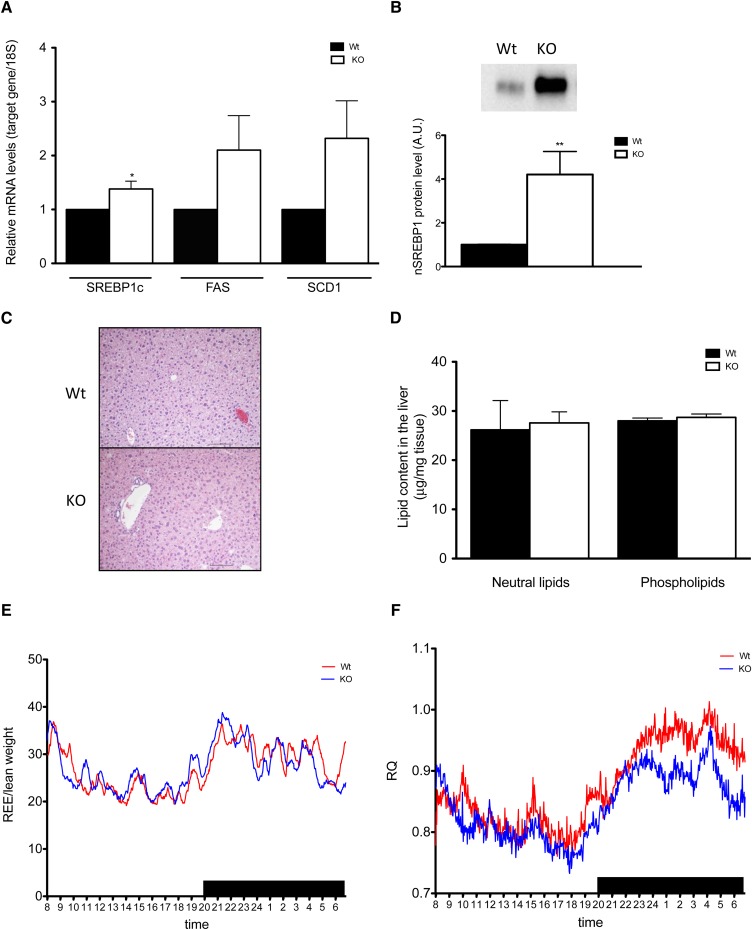
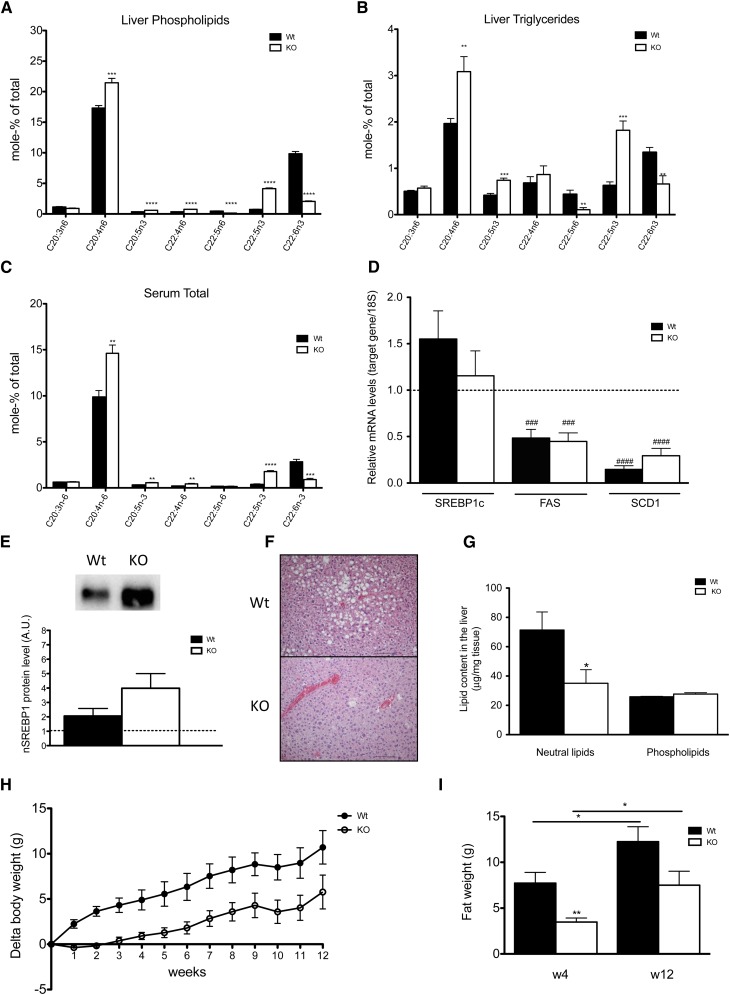
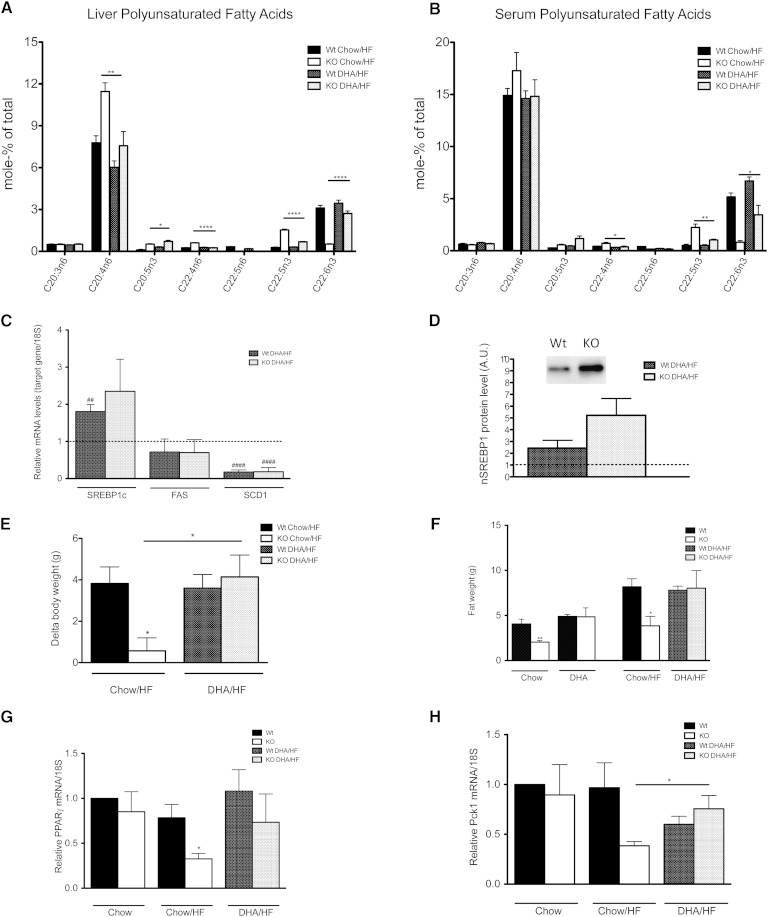
The potential role of endogenously synthesized PUFAs is a highly overlooked area. Elongation of very long-chain fatty acids (ELOVLs) in mammals is catalyzed by the ELOVL enzymes to which the PUFA elongase ELOVL2 belongs. To determine its in vivo function, we have investigated how ablation of ELOVL2, which is highly expressed in liver, affects hepatic lipid composition and function in mice. The Elovl2(-/-) mice displayed substantially decreased levels of 22:6(n-3), DHA, and 22:5(n-6), docosapentaenoic acid (DPA) n-6, and an accumulation of 22:5(n-3) and 22:4(n-6) in both liver and serum, showing that ELOVL2 primarily controls the elongation process of PUFAs with 22 carbons to produce 24-carbon precursors for DHA and DPAn-6 formation in vivo. The impaired PUFA levels positively influenced hepatic levels of the key lipogenic transcriptional regulator sterol-regulatory element binding protein 1c (SREBP-1c), as well as its downstream target genes. Surprisingly, the Elovl2(-/-) mice were resistant to hepatic steatosis and diet-induced weight gain, implying that hepatic DHA synthesis via ELOVL2, in addition to controlling de novo lipogenesis, also regulates lipid storage and fat mass expansion in an SREBP-1c-independent fashion. The changes in fatty acid metabolism were reversed by dietary supplementation with DHA.
Keywords: docosahexaenoic acid; elongase of very long-chain fatty acid 2; fatty acid elongase; liver steatosis; weight gain.
Figures
References
-
- Jump D. B., Clarke S. D. 1999. Regulation of gene expression by dietary fat. Annu. Rev. Nutr. 19: 63–90. - PubMed
-
- Sekiya M., Yahagi N., Matsuzaka T., Najima Y., Nakakuki M., Nagai R., Ishibashi S., Osuga J., Yamada N., Shimano H. 2003. Polyunsaturated fatty acids ameliorate hepatic steatosis in obese mice by SREBP-1 suppression. Hepatology. 38: 1529–1539. - PubMed
-
- González-Périz A., Horrillo R., Ferré N., Gronert K., Dong B., Morán-Salvador E., Titos E., Martínez-Clemente M., López-Parra M., Arroyo V., et al. 2009. Obesity-induced insulin resistance and hepatic steatosis are alleviated by omega-3 fatty acids: a role for resolvins and protectins. FASEB J. 23: 1946–1957. - PMC - PubMed
-
- Burr G. O., Burr M. M. 1973. Nutrition classics from The Journal of Biological Chemistry 82:345–67, 1929. A new deficiency disease produced by the rigid exclusion of fat from the diet. Nutr. Rev. 31: 248–249. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
