

Spiegelzymes® mirror-image hammerhead ribozymes and mirror-image DNAzymes, an alternative to siRNAs and microRNAs to cleave mRNAs in vivo?
- PMID: 24489764
- PMCID: PMC3906056
- DOI: 10.1371/journal.pone.0086673
Spiegelzymes® mirror-image hammerhead ribozymes and mirror-image DNAzymes, an alternative to siRNAs and microRNAs to cleave mRNAs in vivo?
Erratum in
- PLoS One. 2014;9(3):e93798
Abstract
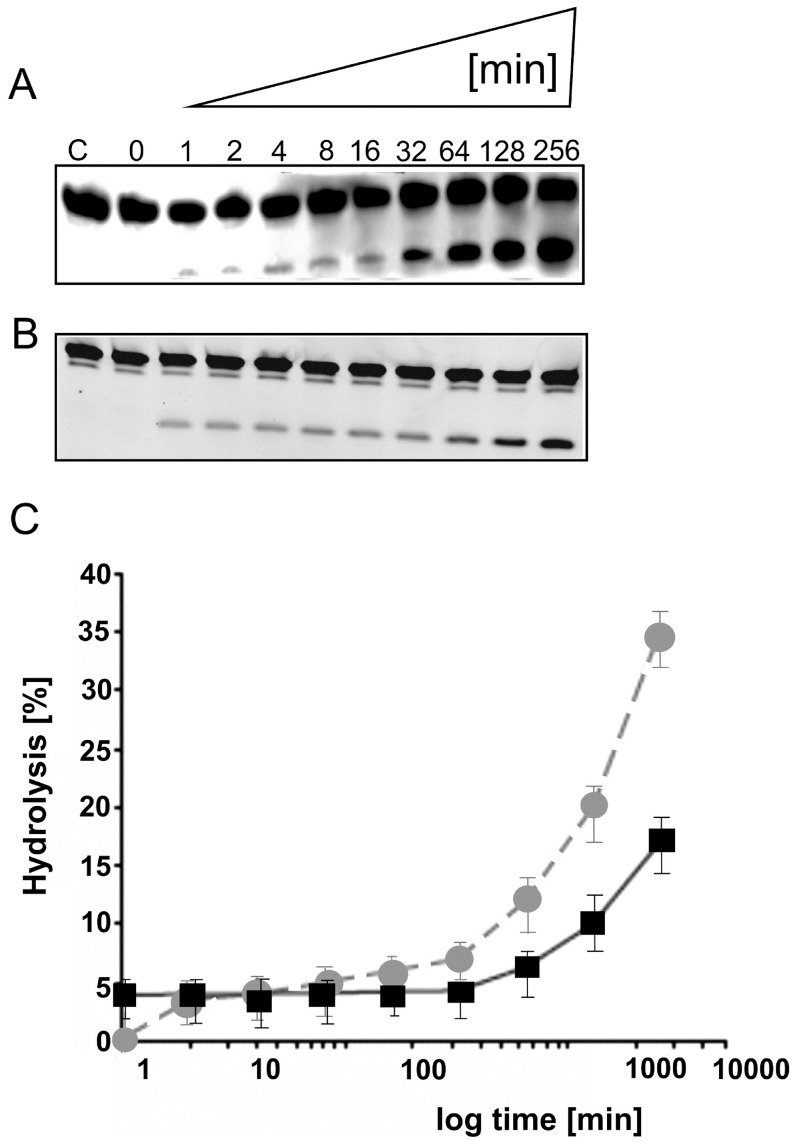
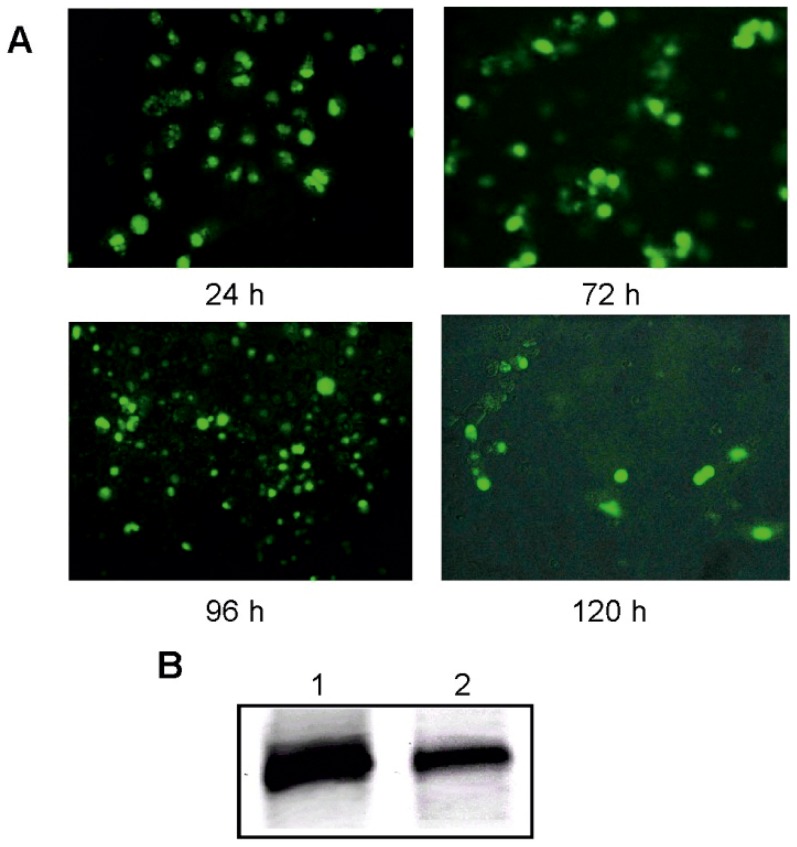
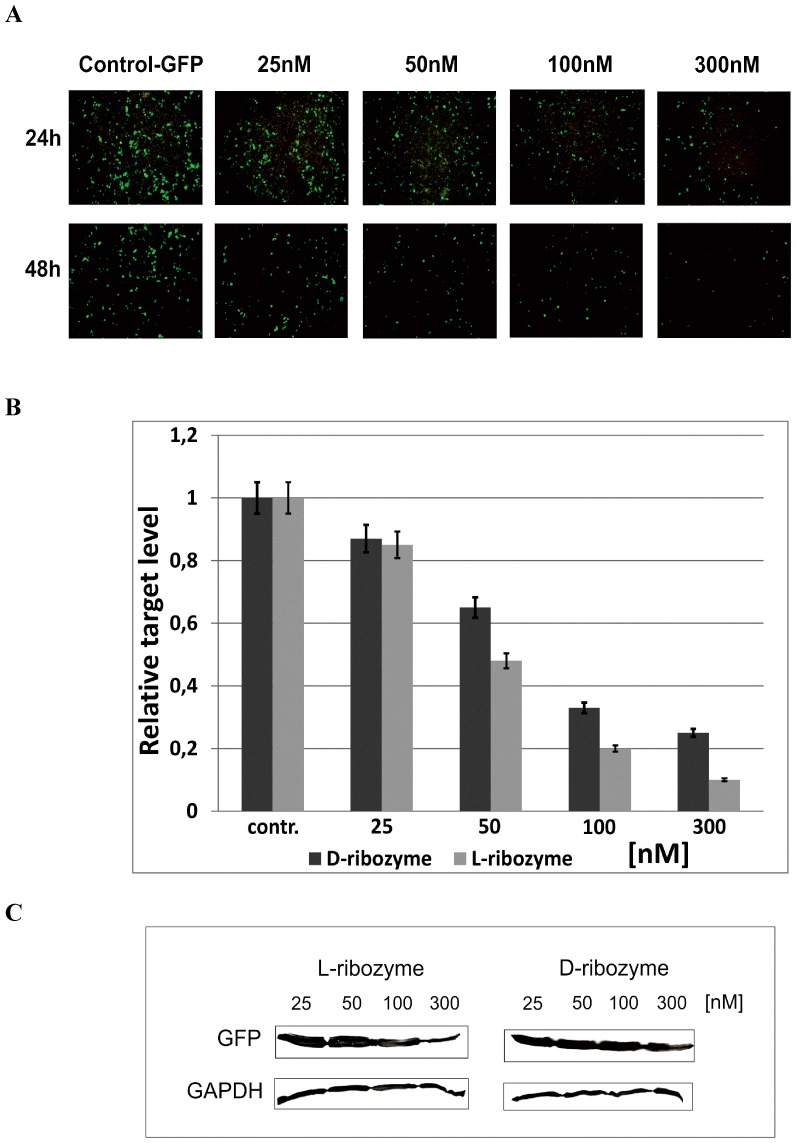
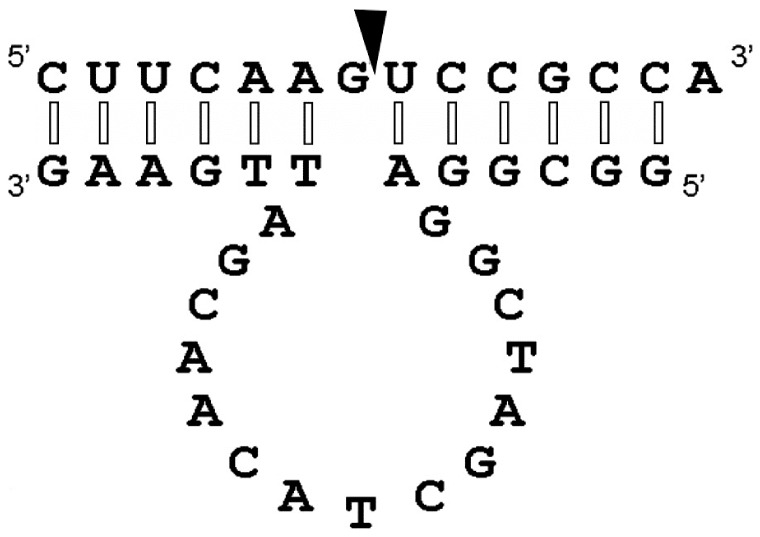
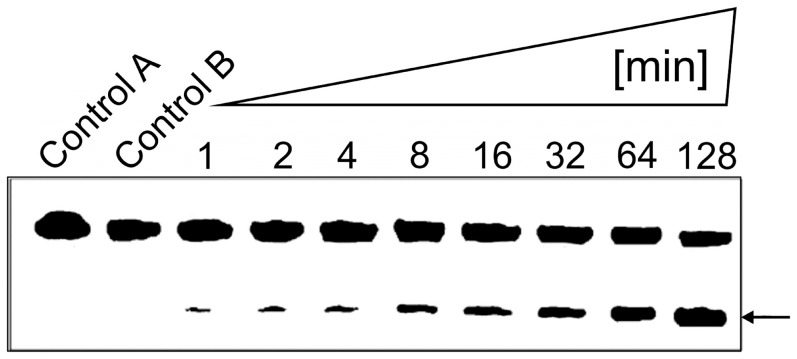
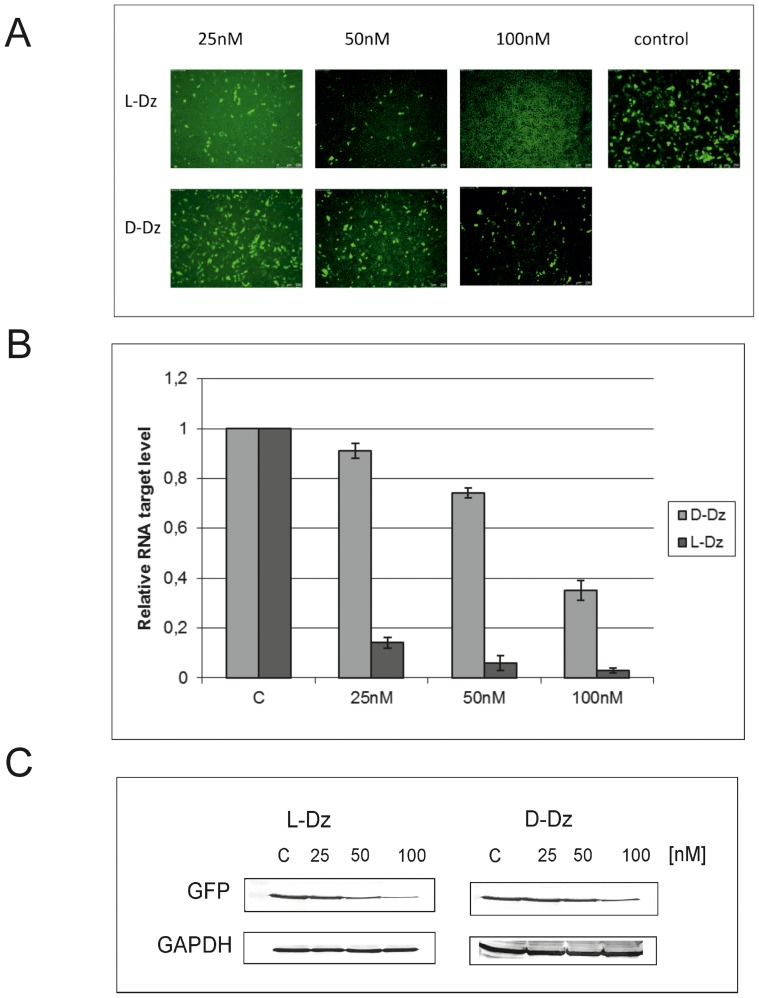
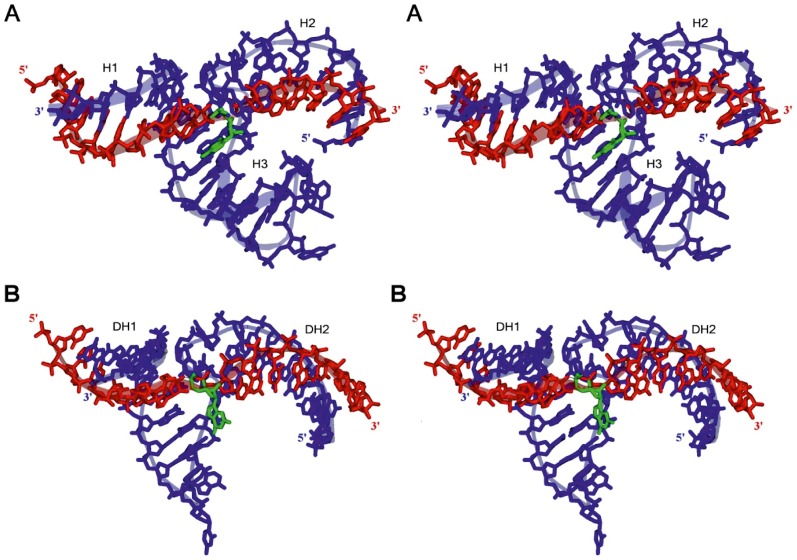
With the discovery of small non-coding RNA (ncRNA) molecules as regulators for cellular processes, it became intriguing to develop technologies by which these regulators can be applied in molecular biology and molecular medicine. The application of ncRNAs has significantly increased our knowledge about the regulation and functions of a number of proteins in the cell. It is surprising that similar successes in applying these small ncRNAs in biotechnology and molecular medicine have so far been very limited. The reasons for these observations may lie in the high complexity in which these RNA regulators function in the cells and problems with their delivery, stability and specificity. Recently, we have described mirror-image hammerhead ribozymes and DNAzymes (Spiegelzymes®) which can sequence-specifically hydrolyse mirror-image nucleic acids, such as our mirror-image aptamers (Spiegelmers) discovered earlier. In this paper, we show for the first time that Spiegelzymes are capable of recognising complementary enantiomeric substrates (D-nucleic acids), and that they efficiently hydrolyse them at submillimolar magnesium concentrations and at physiologically relevant conditions. The Spiegelzymes are very stable in human sera, and do not require any protein factors for their function. They have the additional advantages of being non-toxic and non-immunogenic. The Spiegelzymes can be used for RNA silencing and also as therapeutic and diagnostic tools in medicine. We performed extensive three-dimensional molecular modelling experiments with mirror-image hammerhead ribozymes and DNAzymes interacting with D-RNA targets. We propose a model in which L/D-double helix structures can be formed by natural Watson-Crick base pairs, but where the nucleosides of one of the two strands will occur in an anticlinal conformation. Interestingly enough, the duplexes (L-RNA/D-RNA and L-DNA/D-RNA) in these models can show either right- or left-handedness. This is a very new observation, suggesting that molecular symmetry of enantiomeric nucleic acids is broken down.
Conflict of interest statement
Figures
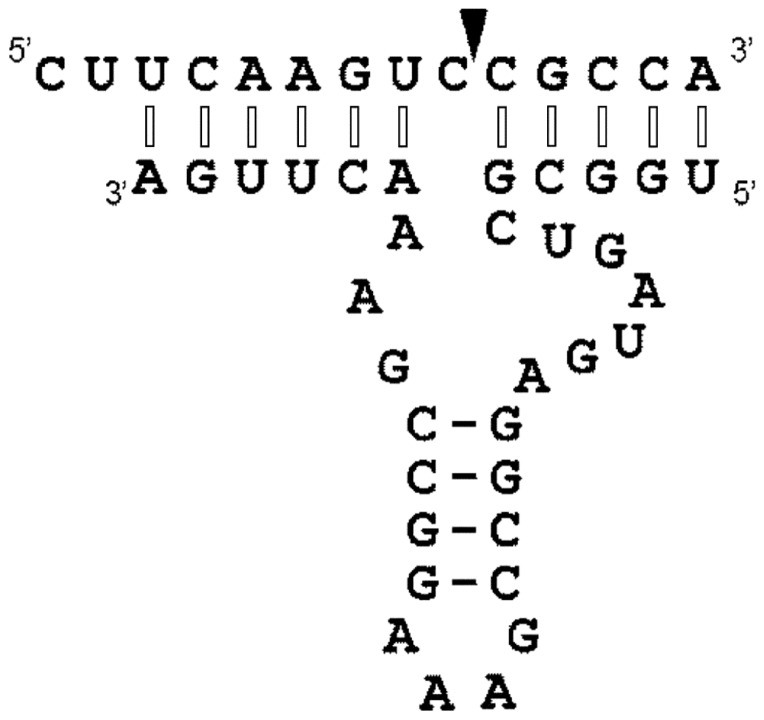
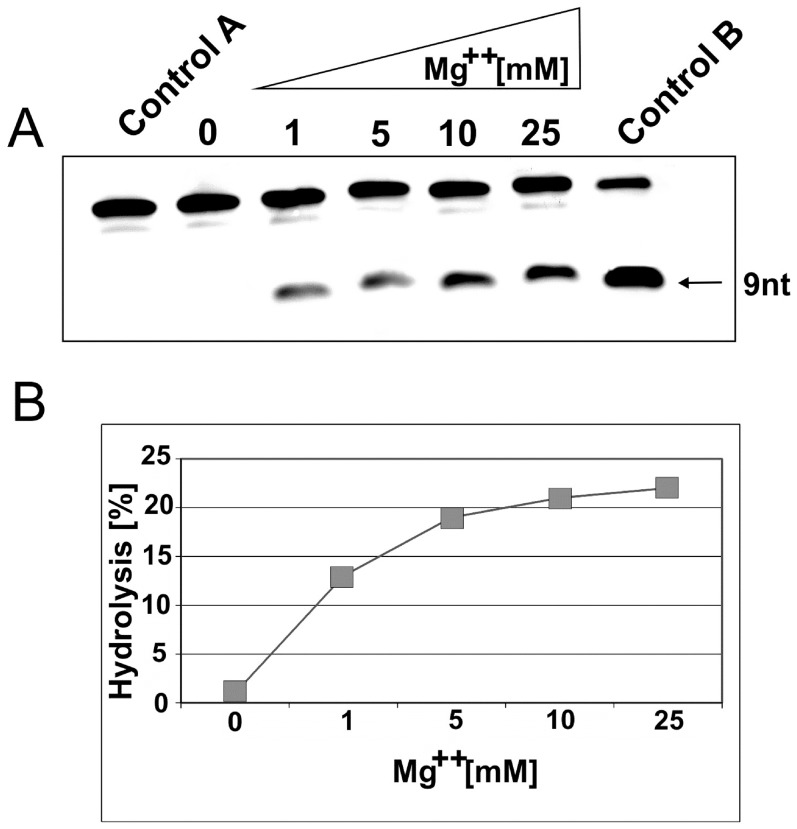
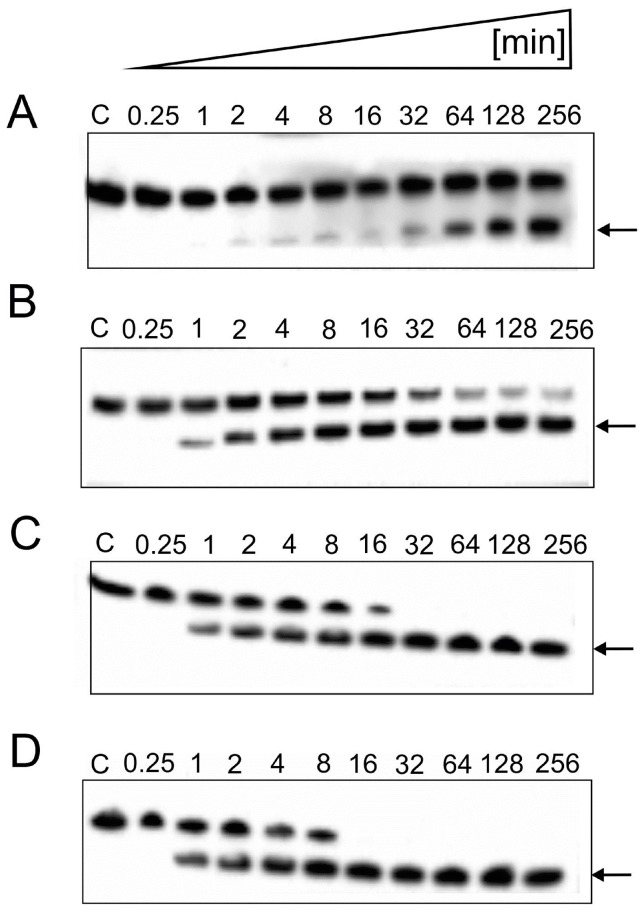
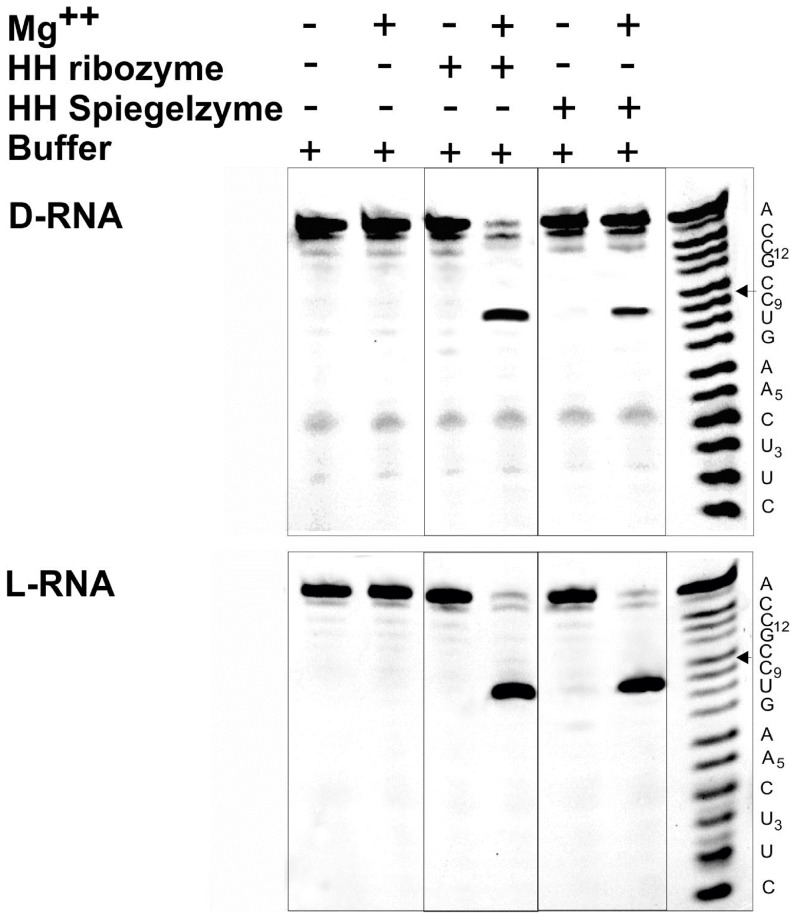

References
-
- Klussmann S, Nolte A, Bald R, Erdmann VA, Fürste JP (1996) Mirror-image RNA that binds D-adenosine. Nat Biotechnol 14: 1112–1115. - PubMed
-
- Nolte A, Klussmann S, Bald R, Erdmann VA, Fürste JP (1996) Mirror-design of L-oligonucleotide ligands binding to L-arginine. Nat Biotechnol 14: 1116–1119. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
