

Rift Valley fever virus incorporates the 78 kDa glycoprotein into virions matured in mosquito C6/36 cells
- PMID: 24489907
- PMCID: PMC3905018
- DOI: 10.1371/journal.pone.0087385
Rift Valley fever virus incorporates the 78 kDa glycoprotein into virions matured in mosquito C6/36 cells
Abstract
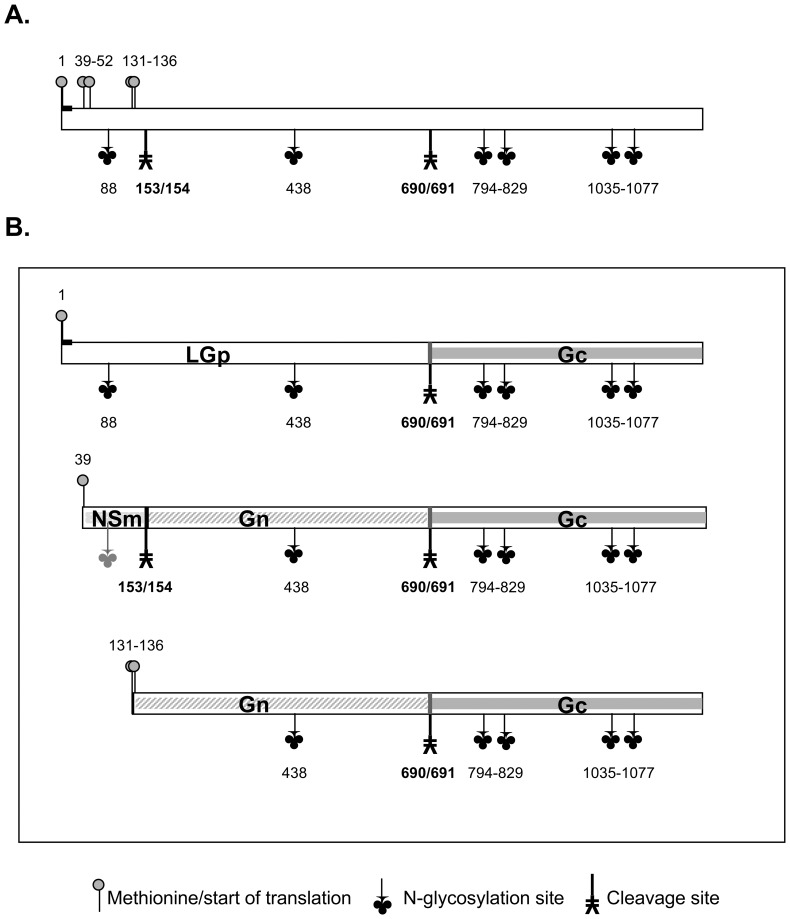
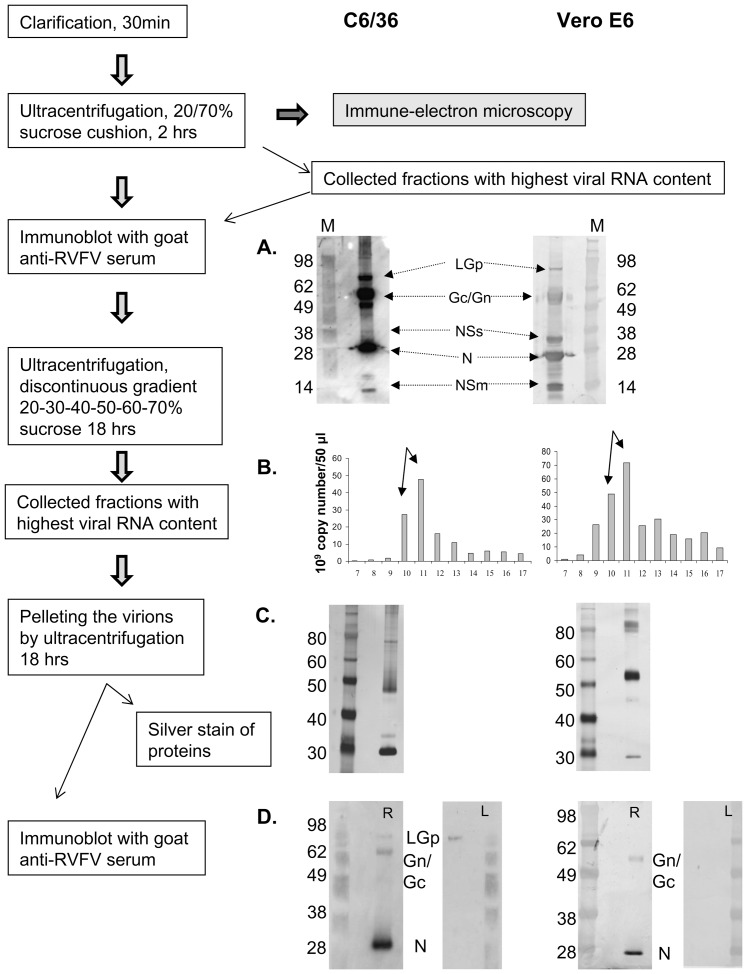
Rift Valley fever virus (RVFV), genus Phlebovirus, family Bunyaviridae is a zoonotic arthropod-borne virus able to transition between distant host species, causing potentially severe disease in humans and ruminants. Viral proteins are encoded by three genomic segments, with the medium M segment coding for four proteins: nonstructural NSm protein, two glycoproteins Gn and Gc and large 78 kDa glycoprotein (LGp) of unknown function. Goat anti-RVFV polyclonal antibody and mouse monoclonal antibody, generated against a polypeptide unique to the LGp within the RVFV proteome, detected this protein in gradient purified RVFV ZH501 virions harvested from mosquito C6/36 cells but not in virions harvested from the mammalian Vero E6 cells. The incorporation of LGp into the mosquito cell line - matured virions was confirmed by immune-electron microscopy. The LGp was incorporated into the virions immediately during the first passage in C6/36 cells of Vero E6 derived virus. Our data indicate that LGp is a structural protein in C6/36 mosquito cell generated virions. The protein may aid the transmission from the mosquitoes to the ruminant host, with a possible role in replication of RVFV in the mosquito host. To our knowledge, this is a first report of different protein composition between virions formed in insect C6/36 versus mammalian Vero E6 cells.
Conflict of interest statement
Figures
References
-
- Collett MS, Purchio AF, Keegan K, Frazier S, Hays W, et al. (1985) Complete nucleotide sequence of the M RNA segment of Rift Valley fever virus. Virology 144: 228–245. - PubMed
-
- Torborg Kakach L, Suzich JA, Collett MS (1989) Rift Valley fever virus M segment: Phlebovirus expression strategy and protein glycosylation. Virology 170: 505–510. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
