

Functional characterization of WalRK: A two-component signal transduction system from Bacillus anthracis
- PMID: 24490131
- PMCID: PMC3907690
- DOI: 10.1016/j.fob.2013.12.005
Functional characterization of WalRK: A two-component signal transduction system from Bacillus anthracis
Abstract
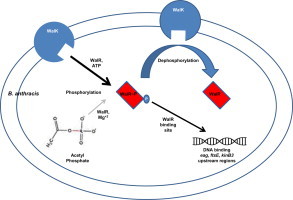
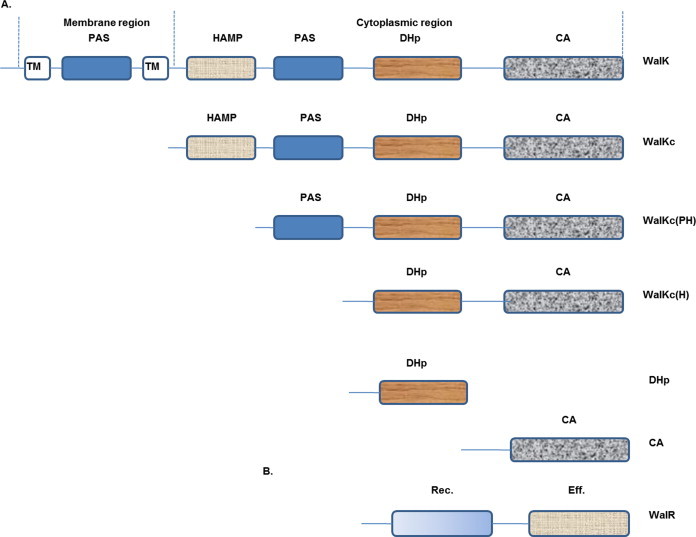
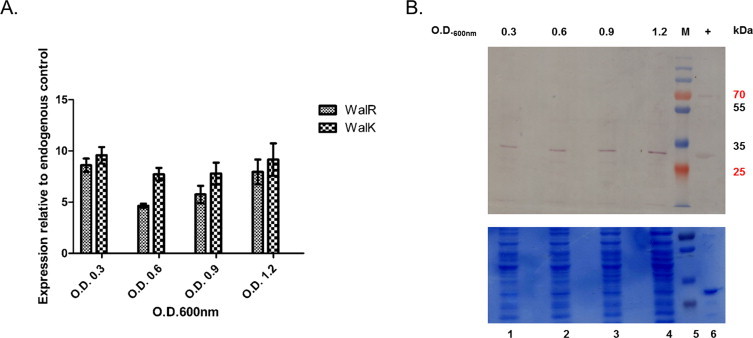
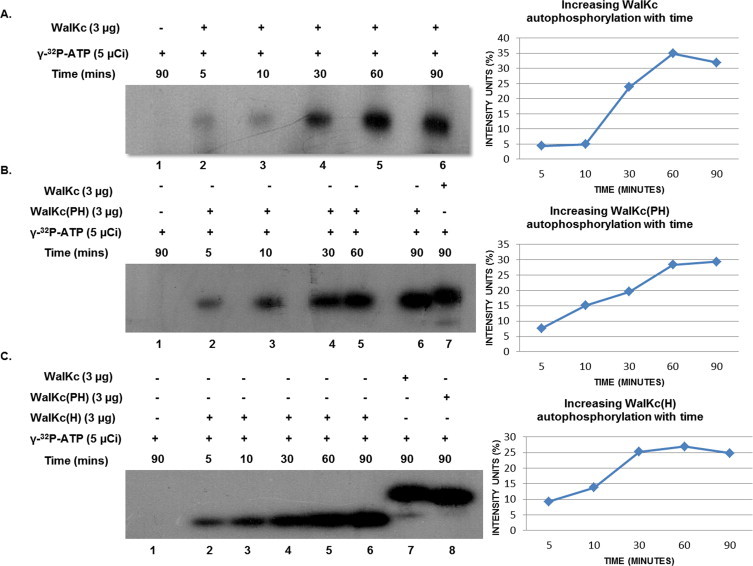
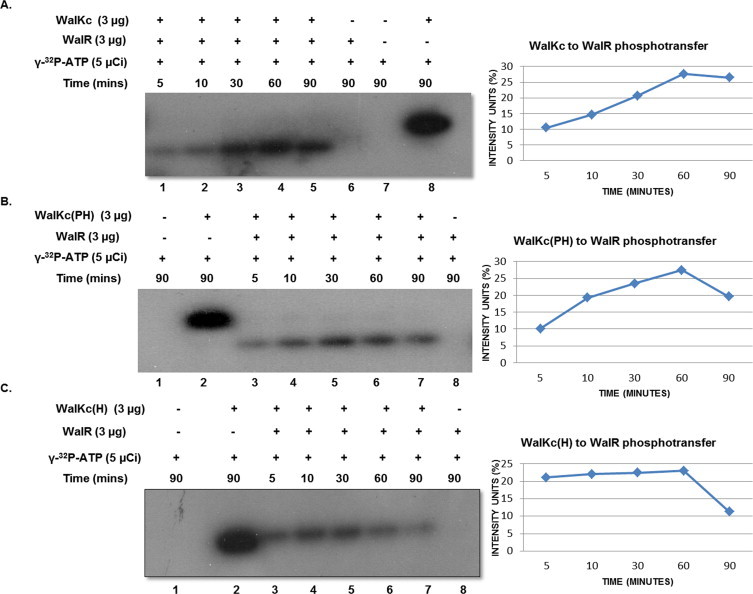
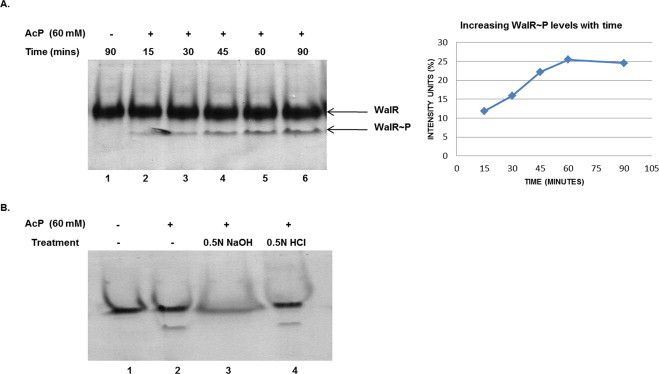
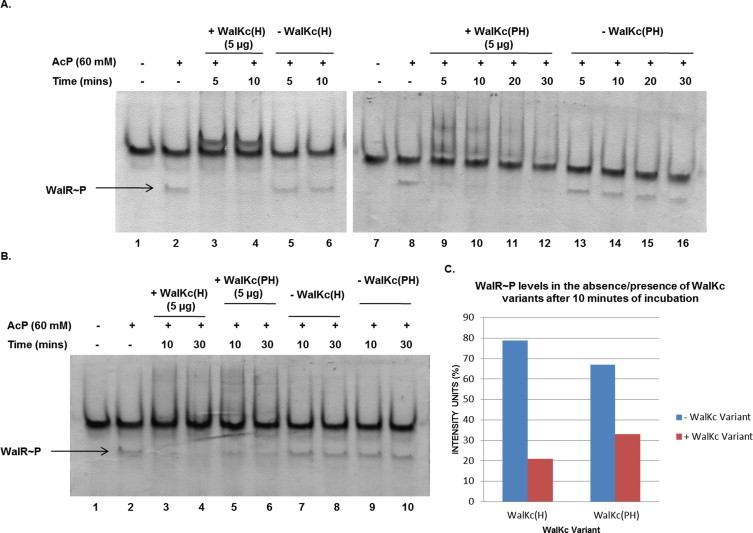
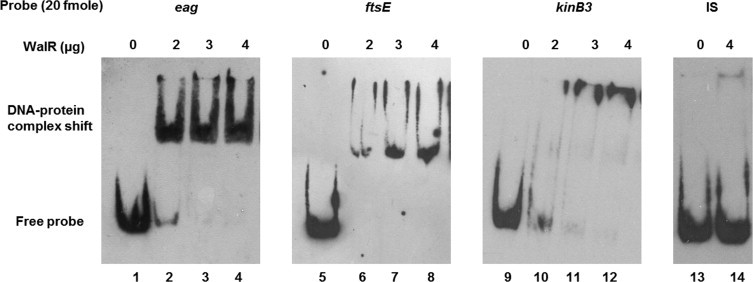
Two-component signal transduction systems (TCS), consisting of a sensor histidine protein kinase and its cognate response regulator, are an important mode of environmental sensing in bacteria. Additionally, they have been found to regulate virulence determinants in several pathogens. Bacillus anthracis, the causative agent of anthrax and a bioterrorism agent, harbours 41 pairs of TCS. However, their role in its pathogenicity has remained largely unexplored. Here, we show that WalRK of B. anthracis forms a functional TCS which exhibits some species-specific functions. Biochemical studies showed that domain variants of WalK, the histidine kinase, exhibit classical properties of autophosphorylation and phosphotransfer to its cognate response regulator WalR. Interestingly, these domain variants also show phosphatase activity towards phosphorylated WalR, thereby making WalK a bifunctional histidine kinase/phosphatase. An in silico regulon determination approach, using a consensus binding sequence from Bacillus subtilis, provided a list of 30 genes that could form a putative WalR regulon in B. anthracis. Further, electrophoretic mobility shift assay was used to show direct binding of purified WalR to the upstream regions of three putative regulon candidates, an S-layer protein EA1, a cell division ABC transporter FtsE and a sporulation histidine kinase KinB3. Our work lends insight into the species-specific functions and mode of action of B. anthracis WalRK.
Keywords: AP, alkaline phosphatase; AcP, acetyl phosphate; Bacillus anthracis; ELISA, enzyme linked immunosorbent assay; EMSA, electrophoretic mobility shift assay; HK, histidine kinase; HRP, horse radish peroxidase; Histidine kinase; RR, response regulator; Regulon; Response regulator; TCS, two-component signal transduction systems; Two-component signal transduction; VicRK; YycFG.
Figures

Similar articles
-
Identification, Functional Characterization and Regulon Prediction of a Novel Two Component System Comprising BAS0540-BAS0541 of Bacillus anthracis.PLoS One. 2016 Jul 8;11(7):e0158895. doi: 10.1371/journal.pone.0158895. eCollection 2016. PLoS One. 2016. PMID: 27392063 Free PMC article.
-
Involvement of WalK (VicK) phosphatase activity in setting WalR (VicR) response regulator phosphorylation level and limiting cross-talk in Streptococcus pneumoniae D39 cells.Mol Microbiol. 2012 Nov;86(3):645-60. doi: 10.1111/mmi.12006. Epub 2012 Sep 27. Mol Microbiol. 2012. PMID: 23013245 Free PMC article.
-
Study on in vivo effects of bacterial histidine kinase inhibitor, Waldiomycin, in Bacillus subtilis and Staphylococcus aureus.J Gen Appl Microbiol. 2015;61(5):177-84. doi: 10.2323/jgam.61.177. J Gen Appl Microbiol. 2015. PMID: 26582287
-
Tearing down the wall: peptidoglycan metabolism and the WalK/WalR (YycG/YycF) essential two-component system.Adv Exp Med Biol. 2008;631:214-28. doi: 10.1007/978-0-387-78885-2_15. Adv Exp Med Biol. 2008. PMID: 18792692 Review.
-
A matter of life and death: cell wall homeostasis and the WalKR (YycGF) essential signal transduction pathway.Mol Microbiol. 2008 Dec;70(6):1307-22. doi: 10.1111/j.1365-2958.2008.06483.x. Epub 2008 Oct 23. Mol Microbiol. 2008. PMID: 19019149 Review.
Cited by
-
Giving a signal: how protein phosphorylation helps Bacillus navigate through different life stages.FEMS Microbiol Rev. 2023 Jul 5;47(4):fuad044. doi: 10.1093/femsre/fuad044. FEMS Microbiol Rev. 2023. PMID: 37533212 Free PMC article. Review.
-
Serine/Threonine Protein Phosphatase PstP of Mycobacterium tuberculosis Is Necessary for Accurate Cell Division and Survival of Pathogen.J Biol Chem. 2016 Nov 11;291(46):24215-24230. doi: 10.1074/jbc.M116.754531. Epub 2016 Oct 7. J Biol Chem. 2016. PMID: 27758870 Free PMC article.
-
RclS Sensor Kinase Modulates Virulence of Pseudomonas capeferrum.Int J Mol Sci. 2022 Jul 26;23(15):8232. doi: 10.3390/ijms23158232. Int J Mol Sci. 2022. PMID: 35897798 Free PMC article.
-
Identification, Functional Characterization and Regulon Prediction of a Novel Two Component System Comprising BAS0540-BAS0541 of Bacillus anthracis.PLoS One. 2016 Jul 8;11(7):e0158895. doi: 10.1371/journal.pone.0158895. eCollection 2016. PLoS One. 2016. PMID: 27392063 Free PMC article.
-
The WalRK Two-Component System Is Essential for Proper Cell Envelope Biogenesis in Clostridioides difficile.J Bacteriol. 2022 Jun 21;204(6):e0012122. doi: 10.1128/jb.00121-22. Epub 2022 May 16. J Bacteriol. 2022. PMID: 35575581 Free PMC article.
References
-
- Mock M, Fouet A. Anthrax. Annu. Rev. Microbiol. 2001;55:647–671. 55/1/647 [pii] - PubMed
-
- Collier RJ, Young JA. Anthrax toxin. Annu. Rev. Cell Dev. Biol. 2003;19:45–70. - PubMed
-
- Chitlaru T, Gat O, Gozlan Y, Ariel N, Shafferman A. Differential proteomic analysis of the Bacillus anthracis secretome: distinct plasmid and chromosome CO2-dependent cross talk mechanisms modulate extracellular proteolytic activities. J. Bacteriol. 2006;188:3551–3571. 188/10/3551 [pii] - PMC - PubMed
-
- Gat O, Mendelson I, Chitlaru T, Ariel N, Altboum Z, Levy H, Weiss S, Grosfeld H, Cohen S, Shafferman A. The solute-binding component of a putative Mn(II) ABC transporter (MntA) is a novel Bacillus anthracis virulence determinant. Mol. Microbiol. 2005;58:533–551. MMI4848 [pii] - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
