

Ebselen induces reactive oxygen species (ROS)-mediated cytotoxicity in Saccharomyces cerevisiae with inhibition of glutamate dehydrogenase being a target
- PMID: 24490132
- PMCID: PMC3907691
- DOI: 10.1016/j.fob.2014.01.002
Ebselen induces reactive oxygen species (ROS)-mediated cytotoxicity in Saccharomyces cerevisiae with inhibition of glutamate dehydrogenase being a target
Erratum in
-
Corrigendum to: Ebselen induces reactive oxygen species (ROS)-mediated cytotoxicity in Saccharomyces cerevisiae with inhibition of glutamate dehydrogenase being a target https://doi.org/10.1016/j.fob.2014.01.002.FEBS Open Bio. 2024 Apr;14(4):721-722. doi: 10.1002/2211-5463.13785. Epub 2024 Mar 1. FEBS Open Bio. 2024. PMID: 38425299 Free PMC article. No abstract available.
Abstract
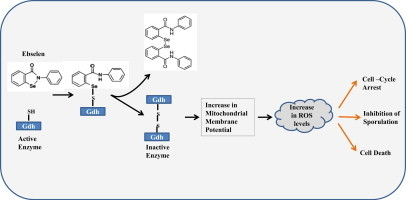
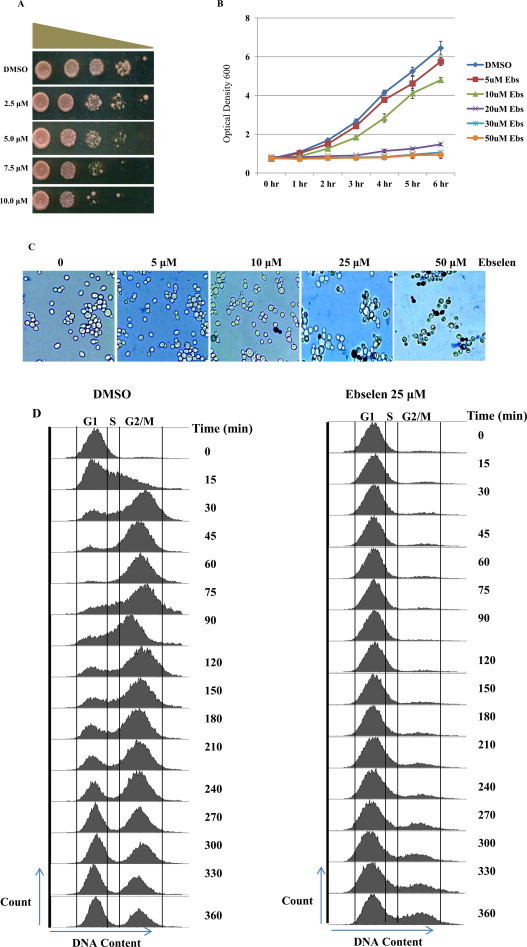
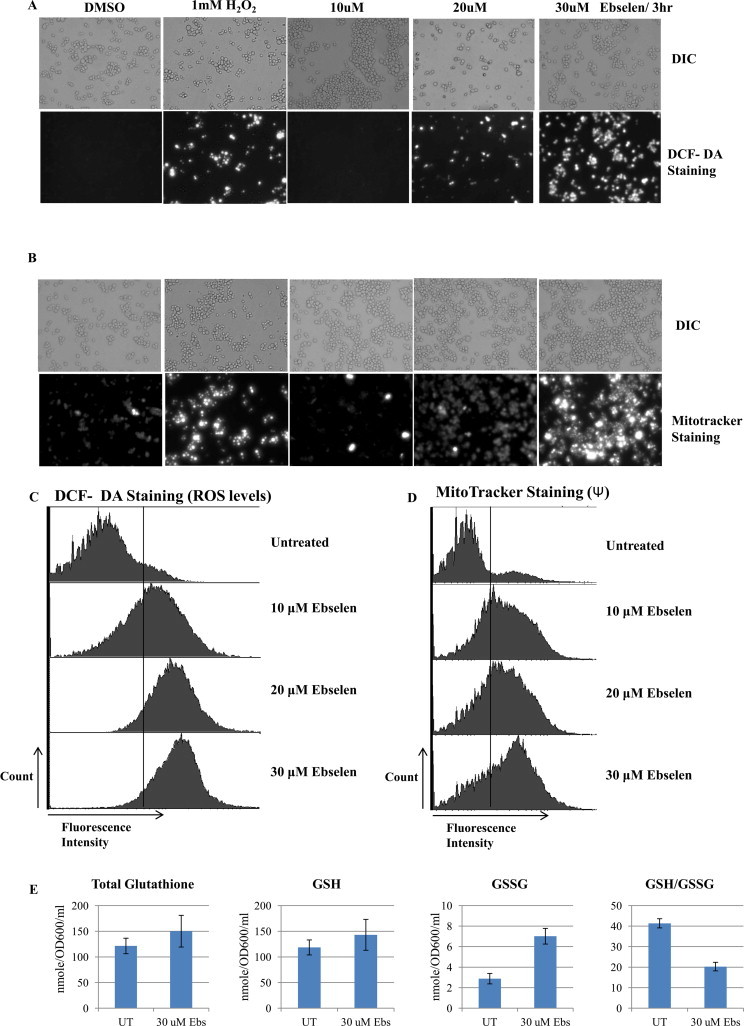
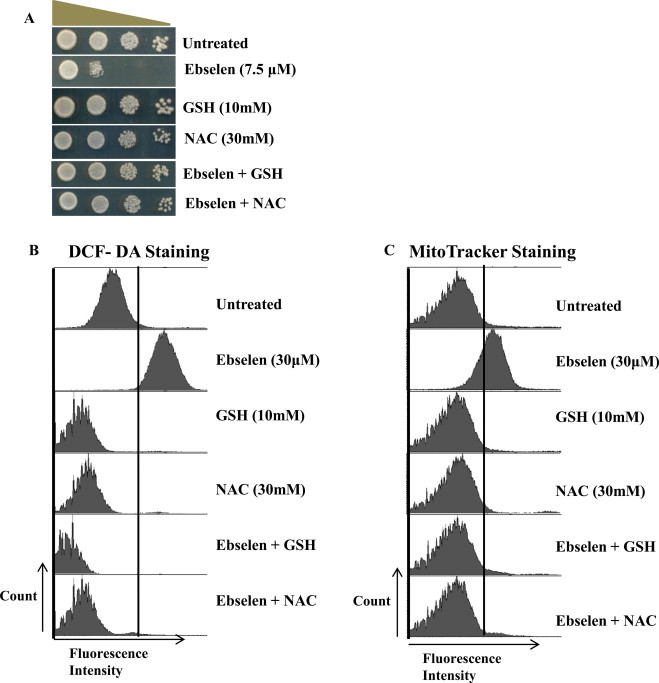
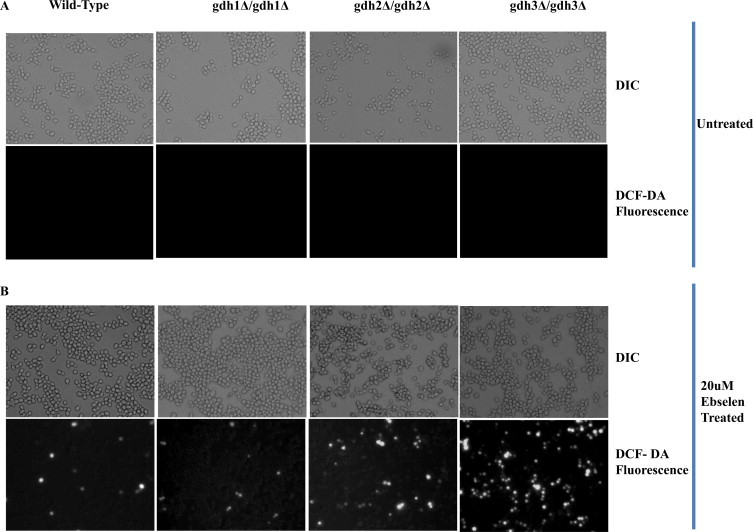
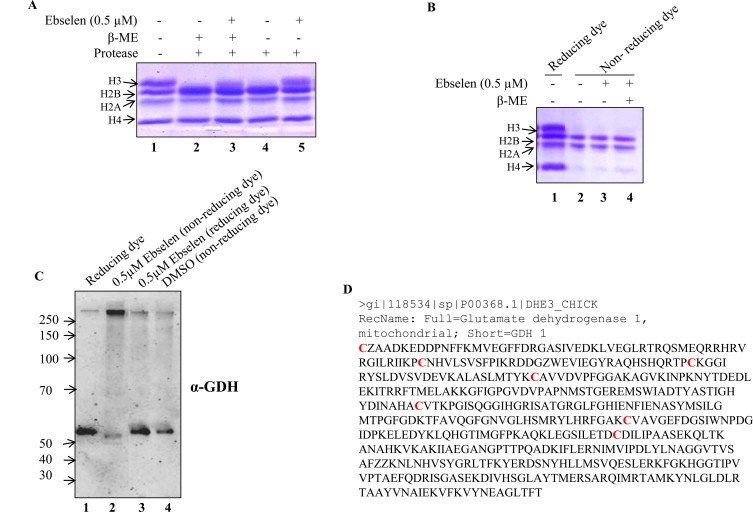
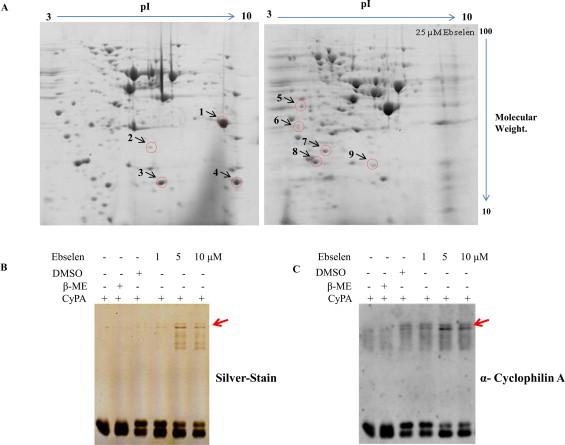
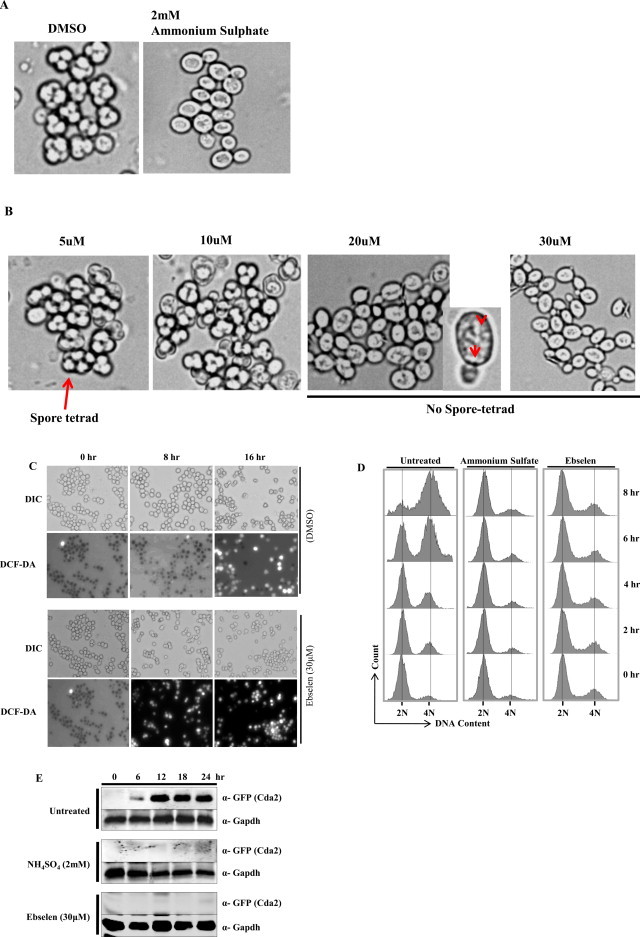
Ebselen is a synthetic, lipid-soluble seleno-organic compound. The high electrophilicity of ebselen enables it to react with multiple cysteine residues of various proteins. Despite extensive research on ebselen, its target molecules and mechanism of action remains less understood. We performed biochemical as well as in vivo experiments employing budding yeast as a model organism to understand the mode of action of ebselen. The growth curve analysis and FACS (florescence activated cell sorting) assays revealed that ebselen exerts growth inhibitory effects on yeast cells by causing a delay in cell cycle progression. We observed that ebselen exposure causes an increase in intracellular ROS levels and mitochondrial membrane potential, and that these effects were reversed by addition of antioxidants such as reduced glutathione (GSH) or N-acetyl-l-cysteine (NAC). Interestingly, a significant increase in ROS levels was noticed in gdh3-deleted cells compared to wild-type cells. Furthermore, we showed that ebselen inhibits GDH function by interacting with its cysteine residues, leading to the formation of inactive hexameric GDH. Two-dimensional gel electrophoresis revealed protein targets of ebselen including CPR1, the yeast homolog of Cyclophilin A. Additionally, ebselen treatment leads to the inhibition of yeast sporulation. These results indicate a novel direct connection between ebselen and redox homeostasis.
Keywords: CypA, Cyclophilin A; DCFH-DA, 2,7-dichlorodihydrofluorescein diacetate; Ebselen; FACS, florescence activated cell sorting; GDH, glutamate dehydrogenase; GSH, glutathione; Glutamate dehydrogenase; Histone clipping; Mitochondrial membrane potential; NAC, N-acetyl-l-cysteine; Ni-NTA, nickel-nitrilotriacetic acid; ROS levels; ROS, reactive oxygen species; SOD, superoxide dismutase; Yeast sporulation.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
