

MiR-21 is an Ngf-modulated microRNA that supports Ngf signaling and regulates neuronal degeneration in PC12 cells
- PMID: 24492999
- PMCID: PMC4019824
- DOI: 10.1007/s12017-014-8292-z
MiR-21 is an Ngf-modulated microRNA that supports Ngf signaling and regulates neuronal degeneration in PC12 cells
Abstract
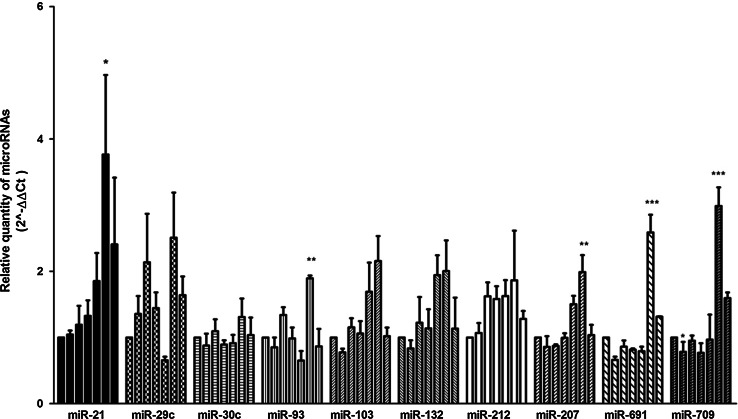
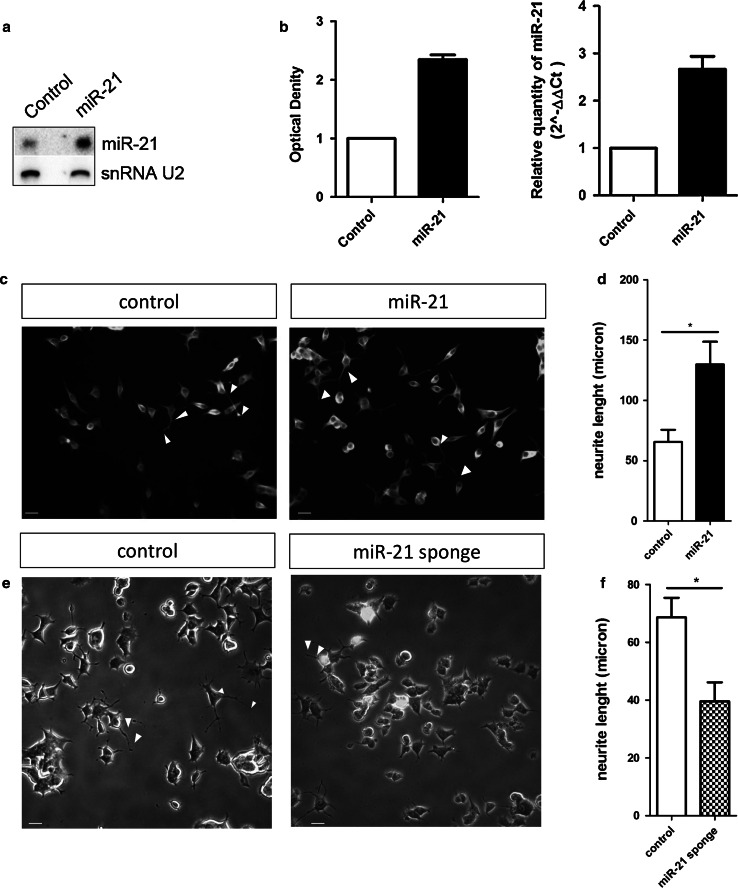
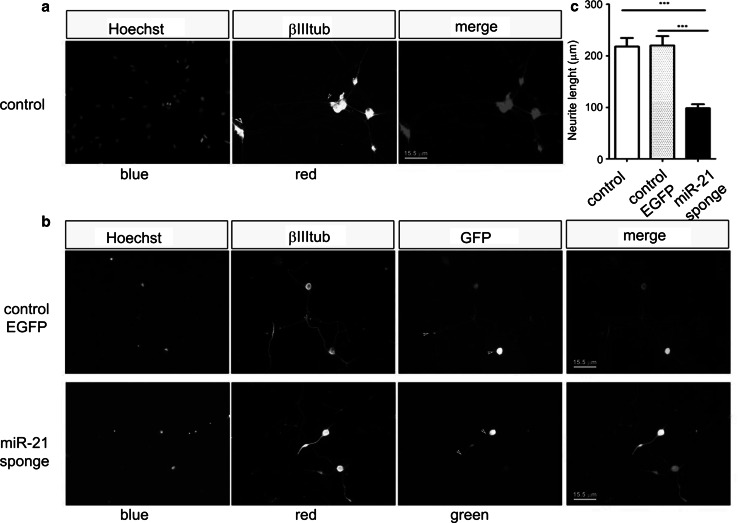
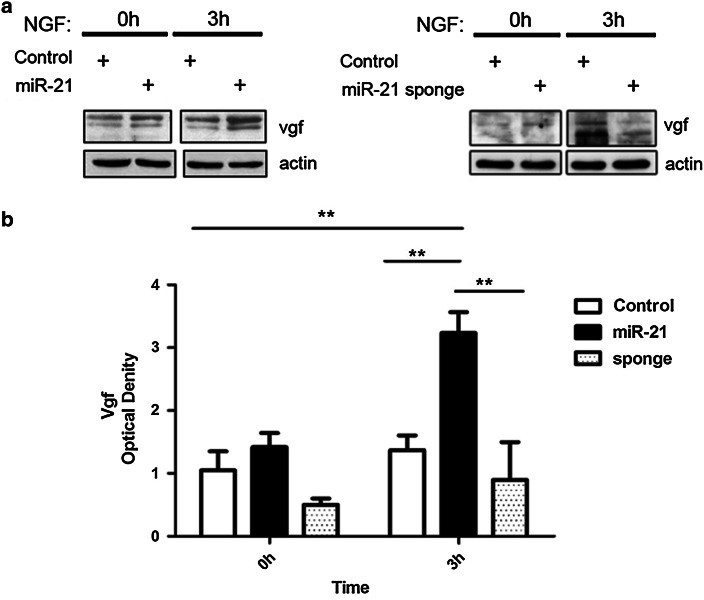
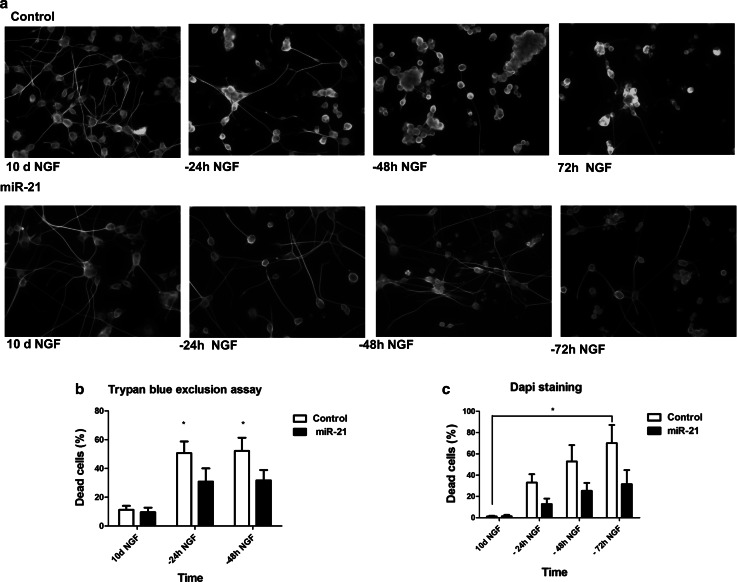
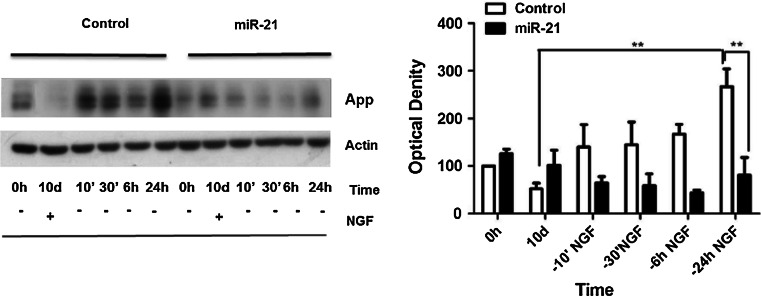
The neurotrophins Ngf, Bdnf, NT-3, NT4-5 have key roles in development, survival, and plasticity of neuronal cells. Their action involves broad gene expression changes at the level of transcription and translation. MicroRNAs (miRs)-small RNA molecules that control gene expression post-transcriptionally-are increasingly implicated in regulating development and plasticity of neural cells. Using PC12 cells as a model system, we show that Ngf modulates changes in expression of a variety of microRNAs, including miRs known to be modulated by neurotrophins-such as the miR-212/132 cluster-and several others, such as miR-21, miR-29c, miR-30c, miR-93, miR-103, miR-207, miR-691, and miR-709. Pathway analysis indicates that Ngf-modulated miRs may regulate many protein components of signaling pathways involved in neuronal development and disease. In particular, we show that miR-21 enhances neurotrophin signaling and controls neuronal differentiation induced by Ngf. Notably, in a situation mimicking neurodegeneration-differentiated neurons deprived of Ngf-this microRNA is able to preserve the neurite network and to support viability of the neurons. These findings uncover a broad role of microRNAs in regulating neurotrophin signaling and suggest that aberrant expression of one or more Ngf-modulated miRs may be involved in neurodegenerative diseases.
Figures

References
-
- Annibali, D., & Nasi, S. (2007). Signaling by neurotrophins. In Reactome. http://www.reactome.org.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
