

Monovalent engagement of the BCR activates ovalbumin-specific transnuclear B cells
- PMID: 24493799
- PMCID: PMC3920557
- DOI: 10.1084/jem.20131603
Monovalent engagement of the BCR activates ovalbumin-specific transnuclear B cells
Abstract
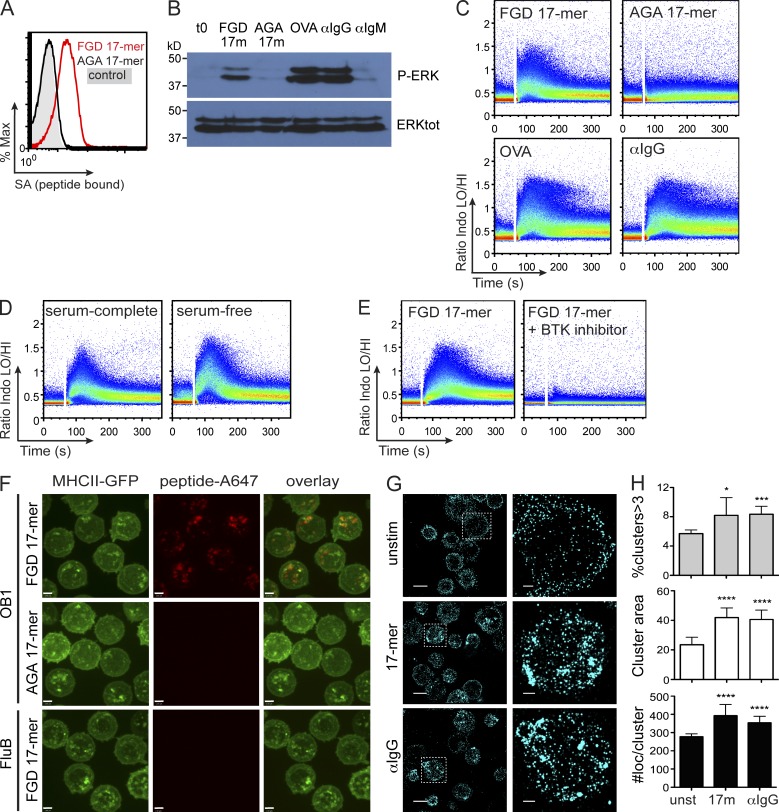
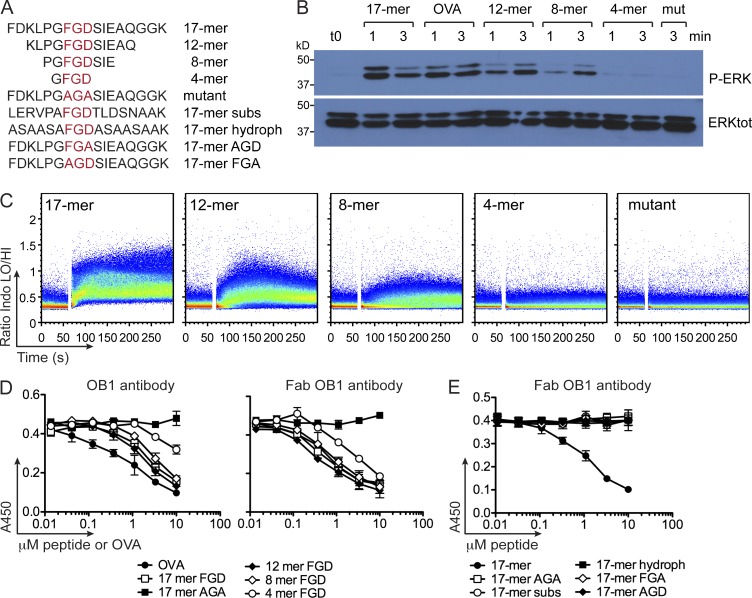
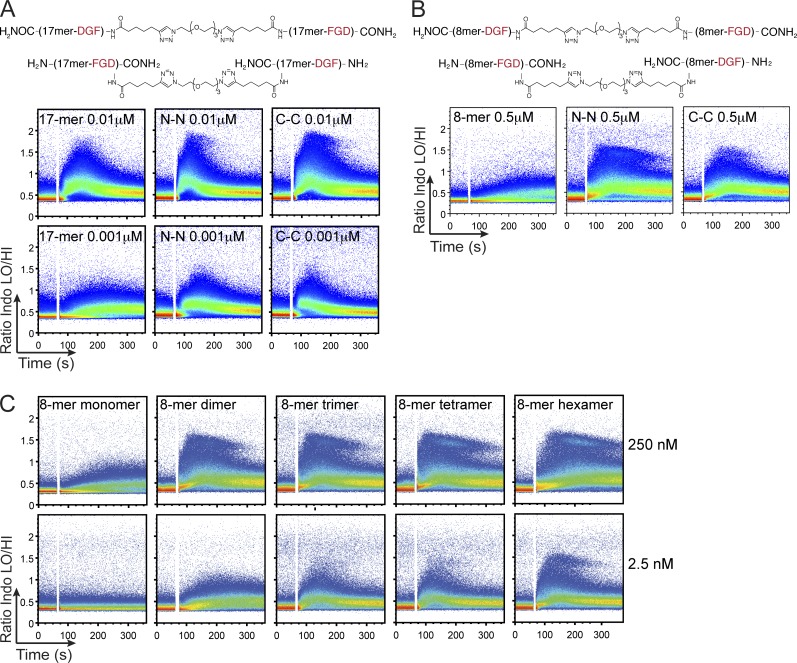
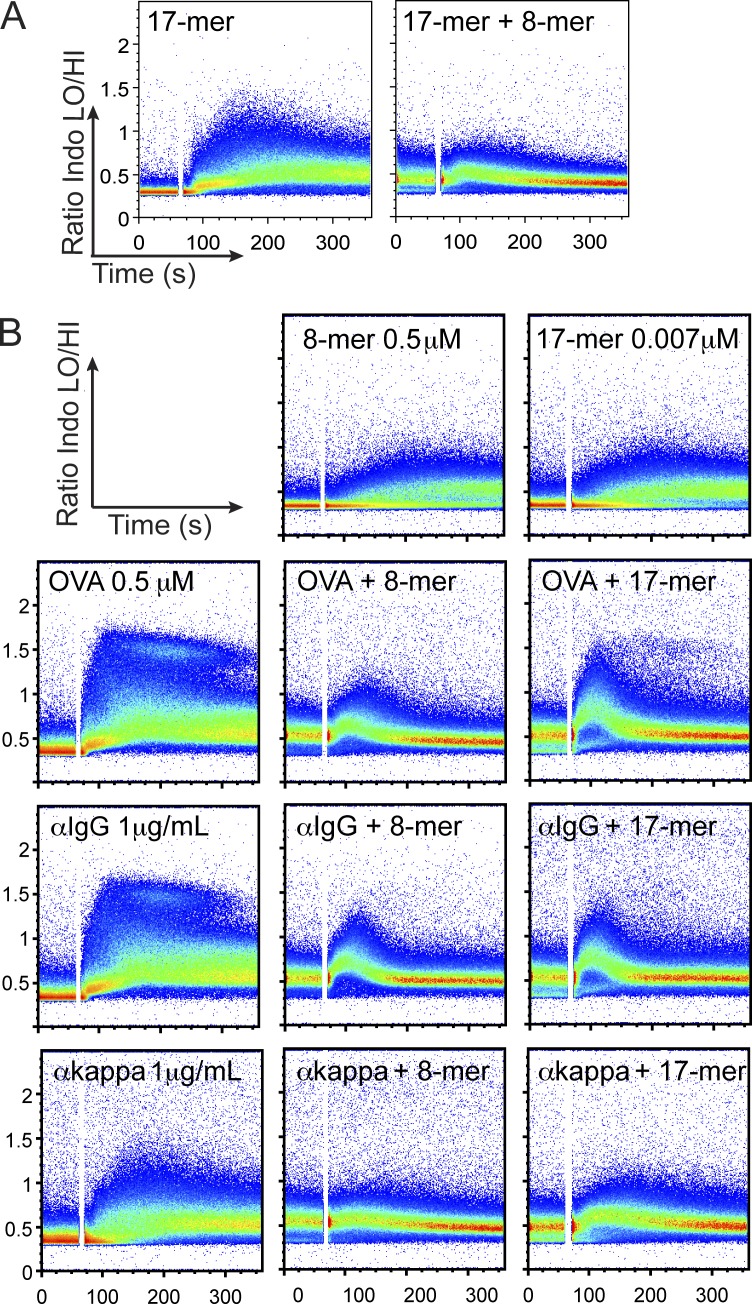
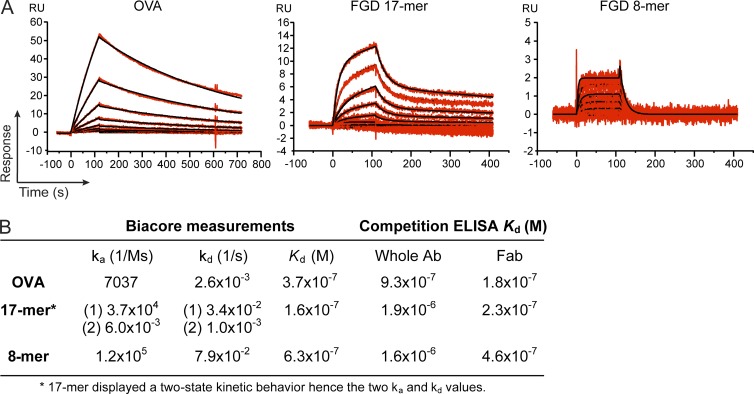
Valency requirements for B cell activation upon antigen encounter are poorly understood. OB1 transnuclear B cells express an IgG1 B cell receptor (BCR) specific for ovalbumin (OVA), the epitope of which can be mimicked using short synthetic peptides to allow antigen-specific engagement of the BCR. By altering length and valency of epitope-bearing synthetic peptides, we examined the properties of ligands required for optimal OB1 B cell activation. Monovalent engagement of the BCR with an epitope-bearing 17-mer synthetic peptide readily activated OB1 B cells. Dimers of the minimal peptide epitope oriented in an N to N configuration were more stimulatory than their C to C counterparts. Although shorter length correlated with less activation, a monomeric 8-mer peptide epitope behaved as a weak agonist that blocked responses to cell-bound peptide antigen, a blockade which could not be reversed by CD40 ligation. The 8-mer not only delivered a suboptimal signal, which blocked subsequent responses to OVA, anti-IgG, and anti-kappa, but also competed for binding with OVA. Our results show that fine-tuning of BCR-ligand recognition can lead to B cell nonresponsiveness, activation, or inhibition.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
