

Complex craniofacial changes in blind cave-dwelling fish are mediated by genetically symmetric and asymmetric loci
- PMID: 24496009
- PMCID: PMC3982692
- DOI: 10.1534/genetics.114.161661
Complex craniofacial changes in blind cave-dwelling fish are mediated by genetically symmetric and asymmetric loci
Abstract
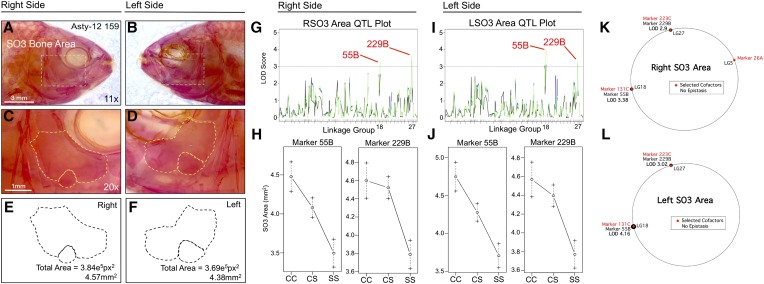
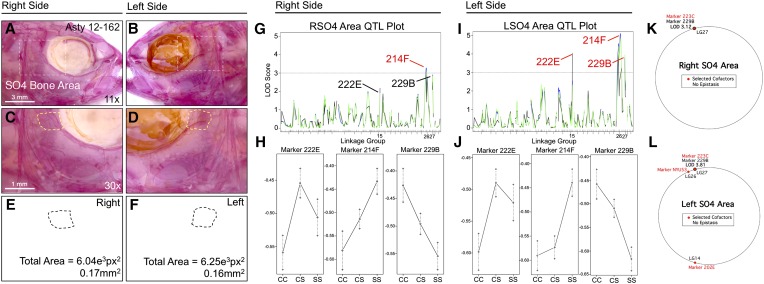
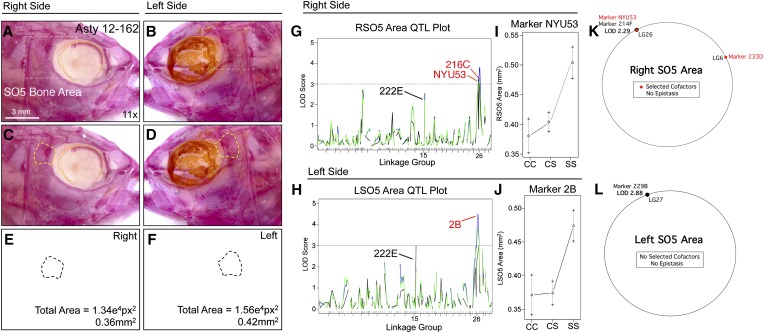
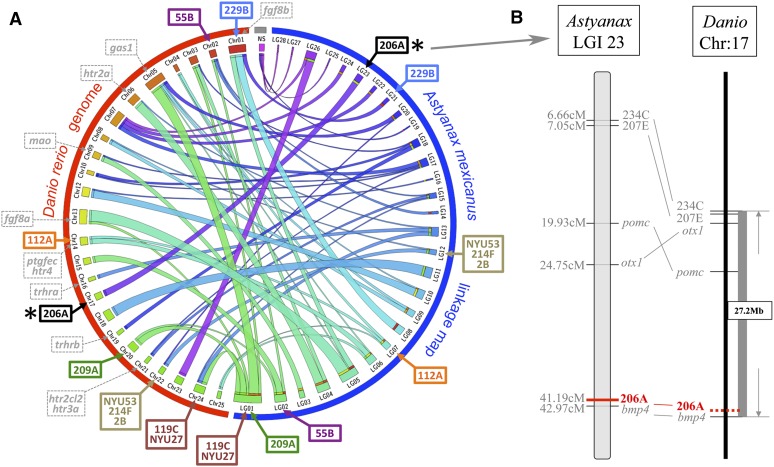
The genetic regulators of regressive craniofacial morphologies are poorly understood. To shed light on this problem, we examined the freshwater fish Astyanax mexicanus, a species with surface-dwelling and multiple independent eyeless cave-dwelling forms. Changes affecting the skull in cavefish include morphological alterations to the intramembranous circumorbital bones encircling the eye. Many of these modifications, however, have evolved separately from eye loss, such as fragmentation of the third suborbital bone. To understand the genetic architecture of these eye-independent craniofacial alterations, we developed and scored 33 phenotypes in the context of an F2 hybrid mapping pedigree bred from Pachón cavefish and surface fish. We discovered several individuals exhibiting dramatic left-right differences in bone formation, such as extensive fragmentation on the right side only. This observation, along with well-known eye size asymmetry in natural cave-dwelling animals, led us to further evaluate left-right genetic differences for the craniofacial complex. We discovered three phenotypes, inclusive of bone fragmentation and fusion, which demonstrated a directional heritable basis only on one side. Interestingly, the overall areas of affected bones were genetically symmetric. Phenotypic effect plots of these novel craniofacial QTL revealed that cave alleles are associated with abnormal conditions such as bony fusion and fragmentation. Moreover, many linked loci overlapped with other cave-associated traits, suggesting regressive craniofacial changes may evolve through linkage or as antagonistic pleiotropic consequences of cave-associated adaptations. These novel findings illuminate significant craniofacial changes accompanying evolution in complete darkness and reveal complex changes to the skull differentially influenced by genetic changes affecting the left and right sides.
Keywords: Astyanax; circumorbital bone series; quantitative trait locus analysis; regressive phenotypic evolution; troglomorphy.
Figures
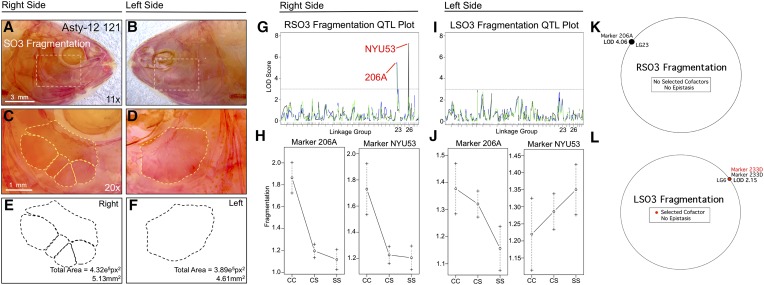
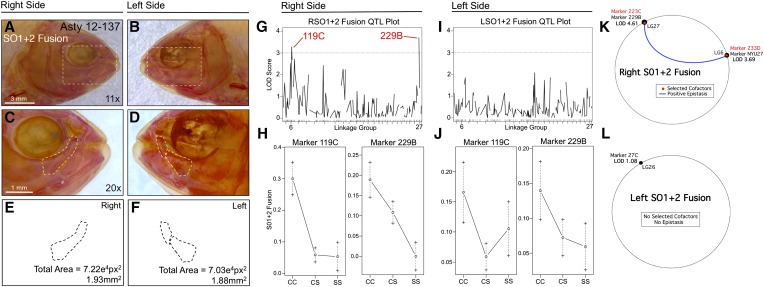
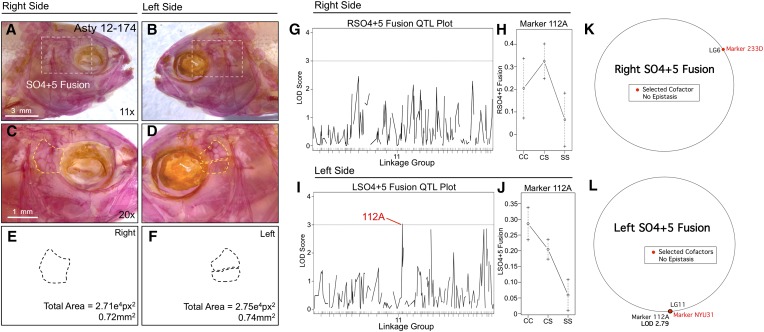
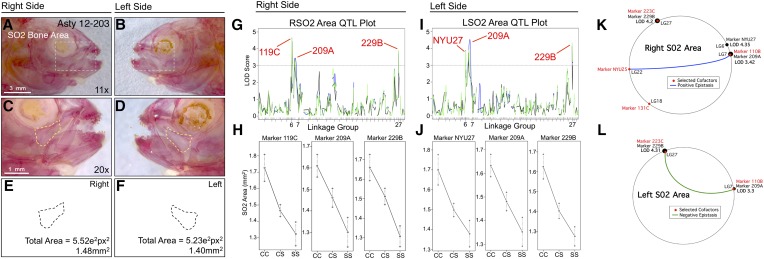
References
-
- Abzhanov A., Protas M., Grant B. R., Grant P. R., Tabin C. J., 2004. Bmp4 and morphological variation of beaks in Darwin’s finches. Science 305: 1462–1465. - PubMed
-
- Abzhanov A., Kuo W. P., Hartmann C., Grant B. R., Grant P. R., et al. , 2006. The calmodulin pathway and evolution of elongated beak morphology in Darwin’s finches. Nature 442: 563–567. - PubMed
-
- Albertson R. C., Kocher T. D., 2006. Genetic and developmental basis of cichlid trophic diversity. Heredity 97: 211–221. - PubMed
-
- Albertson R. C., Yelick P. C., 2005. Roles for fgf8 signaling in left–right patterning of the visceral organs and craniofacial skeleton. Dev. Biol. 283: 310–321. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
