

Macroglobulin complement-related encodes a protein required for septate junction organization and paracellular barrier function in Drosophila
- PMID: 24496625
- PMCID: PMC3912832
- DOI: 10.1242/dev.102152
Macroglobulin complement-related encodes a protein required for septate junction organization and paracellular barrier function in Drosophila
Abstract
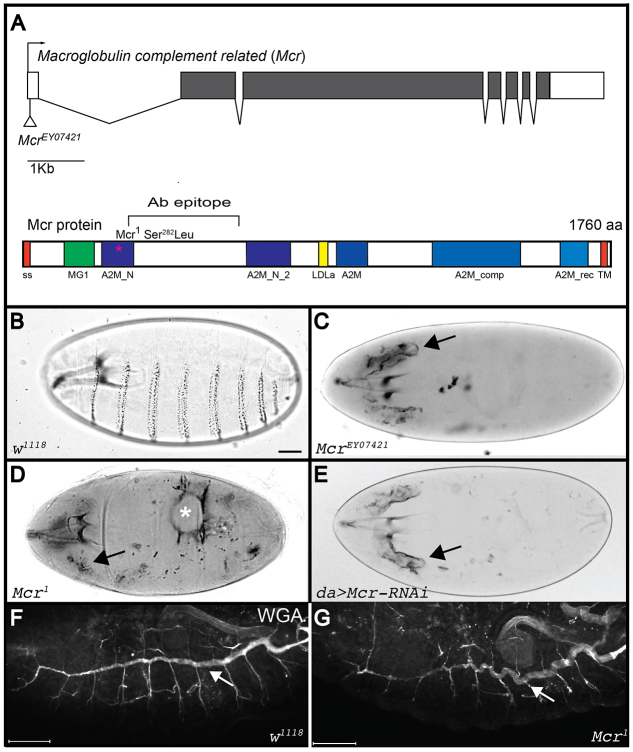
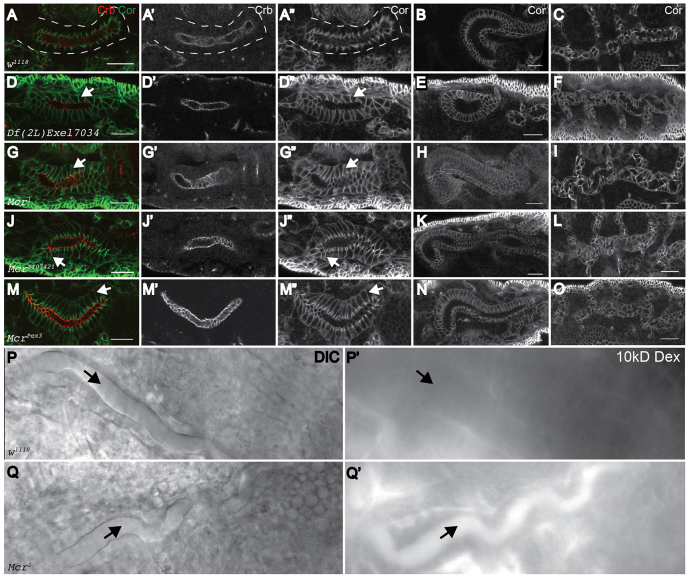
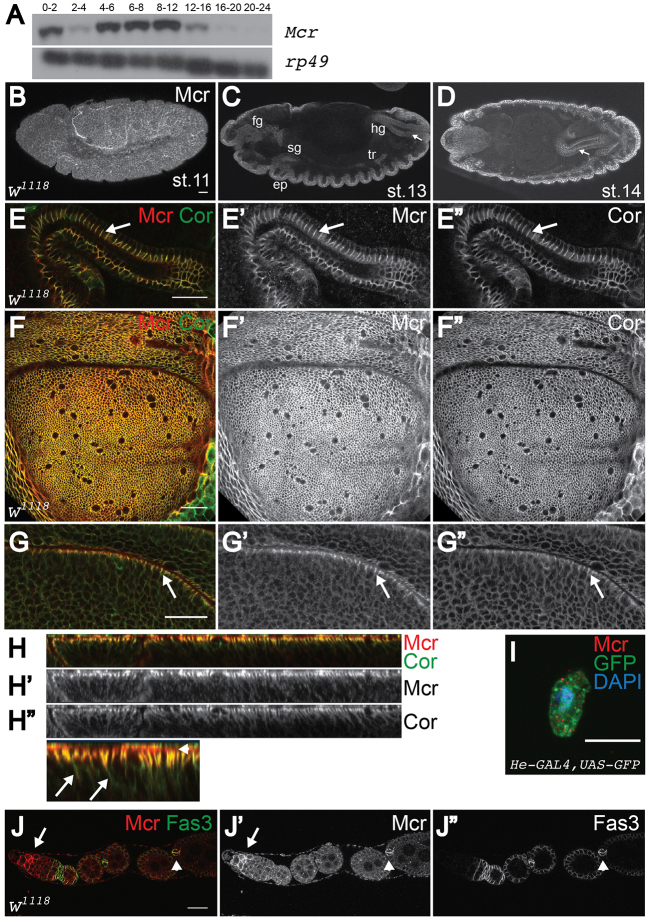
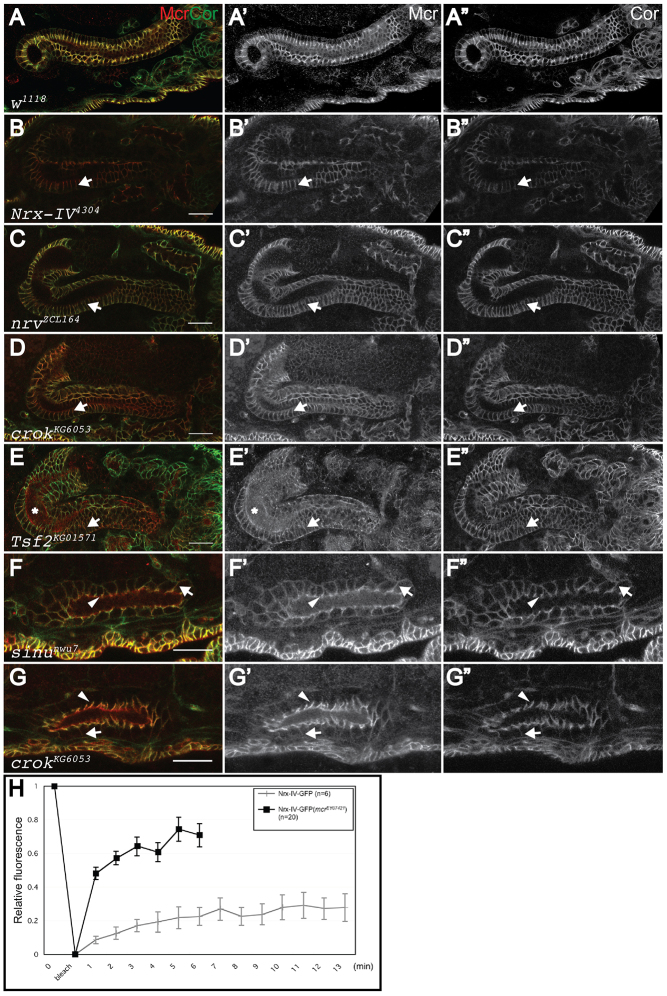
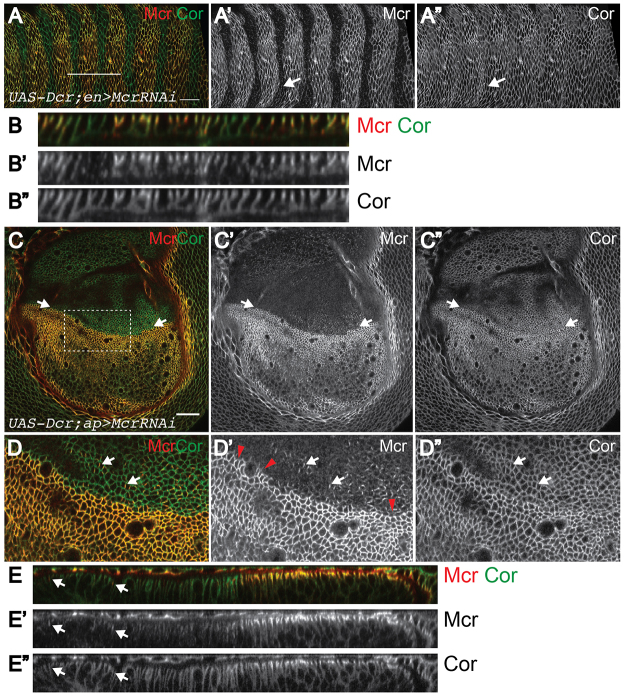
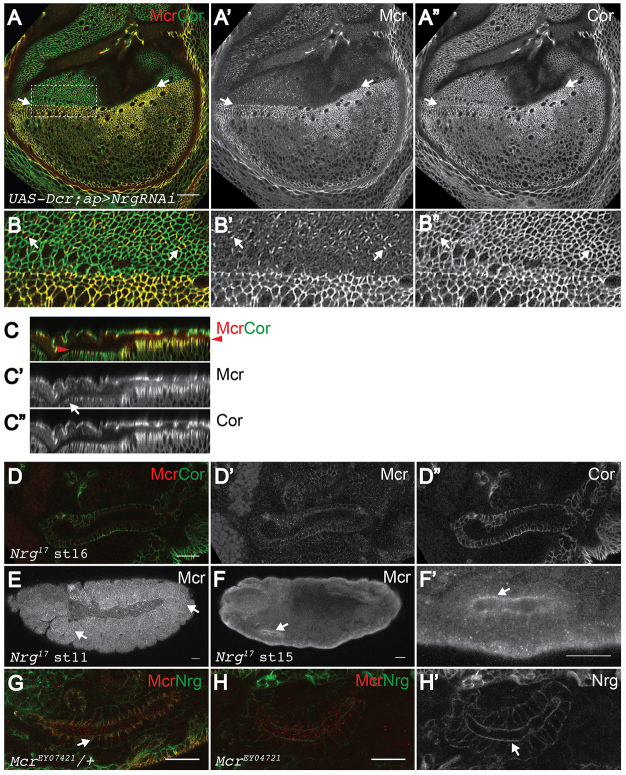
Polarized epithelia play crucial roles as barriers to the outside environment and enable the formation of specialized compartments for organs to carry out essential functions. Barrier functions are mediated by cellular junctions that line the lateral plasma membrane between cells, principally tight junctions in vertebrates and septate junctions (SJs) in invertebrates. Over the last two decades, more than 20 genes have been identified that function in SJ biogenesis in Drosophila, including those that encode core structural components of the junction such as Neurexin IV, Coracle and several claudins, as well as proteins that facilitate the trafficking of SJ proteins during their assembly. Here we demonstrate that Macroglobulin complement-related (Mcr), a gene previously implicated in innate immunity, plays an essential role during embryonic development in SJ organization and function. We show that Mcr colocalizes with other SJ proteins in mature ectodermally derived epithelial cells, that it shows interdependence with other SJ proteins for SJ localization, and that Mcr mutant epithelia fail to form an effective paracellular barrier. Tissue-specific RNA interference further demonstrates that Mcr is required cell-autonomously for SJ organization. Finally, we show a unique interdependence between Mcr and Nrg for SJ localization that provides new insights into the organization of the SJ. Together, these studies demonstrate that Mcr is a core component of epithelial SJs and also highlight an interesting relationship between innate immunity and epithelial barrier functions.
Keywords: Epithelia; Innate immunity; Septate junction.
Figures
Similar articles
-
The transmembrane protein Macroglobulin complement-related is essential for septate junction formation and epithelial barrier function in Drosophila.Development. 2014 Feb;141(4):899-908. doi: 10.1242/dev.102160. Development. 2014. PMID: 24496626
-
Analysis of protein dynamics within the septate junction reveals a highly stable core protein complex that does not include the basolateral polarity protein Discs large.J Cell Sci. 2011 Aug 15;124(Pt 16):2861-71. doi: 10.1242/jcs.087700. J Cell Sci. 2011. PMID: 21807950 Free PMC article.
-
The lateral mobility of cell adhesion molecules is highly restricted at septate junctions in Drosophila.BMC Cell Biol. 2008 Jul 18;9:38. doi: 10.1186/1471-2121-9-38. BMC Cell Biol. 2008. PMID: 18638384 Free PMC article.
-
Molecular organization and function of invertebrate occluding junctions.Semin Cell Dev Biol. 2014 Dec;36:186-93. doi: 10.1016/j.semcdb.2014.09.009. Epub 2014 Sep 17. Semin Cell Dev Biol. 2014. PMID: 25239398 Review.
-
The Drosophila septate junctions beyond barrier function: Review of the literature, prediction of human orthologs of the SJ-related proteins and identification of protein domain families.Acta Physiol (Oxf). 2021 Jan;231(1):e13527. doi: 10.1111/apha.13527. Epub 2020 Aug 5. Acta Physiol (Oxf). 2021. PMID: 32603029 Review.
Cited by
-
Pasiflora proteins are novel core components of the septate junction.Development. 2015 Sep 1;142(17):3046-57. doi: 10.1242/dev.119412. Development. 2015. PMID: 26329602 Free PMC article.
-
Expanding the Junction: New Insights into Non-Occluding Roles for Septate Junction Proteins during Development.J Dev Biol. 2021 Mar 21;9(1):11. doi: 10.3390/jdb9010011. J Dev Biol. 2021. PMID: 33801162 Free PMC article. Review.
-
Tsetse fly tolerance to T. brucei infection: transcriptome analysis of trypanosome-associated changes in the tsetse fly salivary gland.BMC Genomics. 2016 Nov 25;17(1):971. doi: 10.1186/s12864-016-3283-0. BMC Genomics. 2016. PMID: 27884110 Free PMC article.
-
The Drosophila tricellular junction protein Gliotactin regulates its own mRNA levels through BMP-mediated induction of miR-184.J Cell Sci. 2016 Apr 1;129(7):1477-89. doi: 10.1242/jcs.178608. Epub 2016 Feb 16. J Cell Sci. 2016. PMID: 26906422 Free PMC article.
-
BgTEP: An Antiprotease Involved in Innate Immune Sensing in Biomphalaria glabrata.Front Immunol. 2018 May 29;9:1206. doi: 10.3389/fimmu.2018.01206. eCollection 2018. Front Immunol. 2018. PMID: 29899746 Free PMC article.
References
-
- Armstrong P. B., Quigley J. P. (1999). Alpha2-macroglobulin: an evolutionarily conserved arm of the innate immune system. Dev. Comp. Immunol. 23, 375–390 - PubMed
-
- Baumgartner S., Littleton J. T., Broadie K., Bhat M. A., Harbecke R., Lengyel J. A., Chiquet-Ehrismann R., Prokop A., Bellen H. J. (1996). A Drosophila neurexin is required for septate junction and blood-nerve barrier formation and function. Cell 87, 1059–1068 - PubMed
-
- Behr M., Riedel D., Schuh R. (2003). The claudin-like megatrachea is essential in septate junctions for the epithelial barrier function in Drosophila. Dev. Cell 5, 611–620 - PubMed
-
- Bieber A. J., Snow P. M., Hortsch M., Patel N. H., Jacobs J. R., Traquina Z. R., Schilling J., Goodman C. S. (1989). Drosophila neuroglian: a member of the immunoglobulin superfamily with extensive homology to the vertebrate neural adhesion molecule L1. Cell 59, 447–460 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
