

Integrated redox proteomics and metabolomics of mitochondria to identify mechanisms of cd toxicity
- PMID: 24496640
- PMCID: PMC4007108
- DOI: 10.1093/toxsci/kfu018
Integrated redox proteomics and metabolomics of mitochondria to identify mechanisms of cd toxicity
Abstract
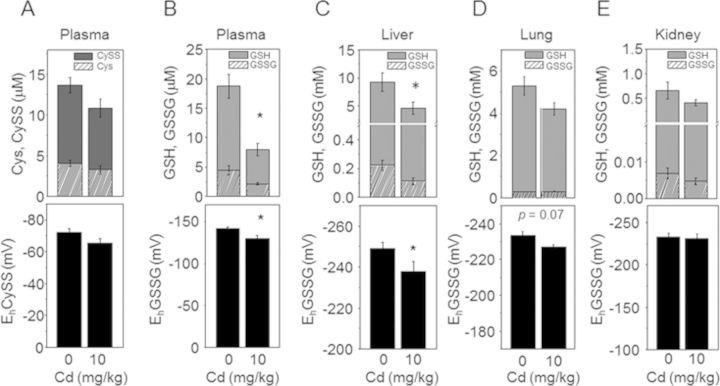
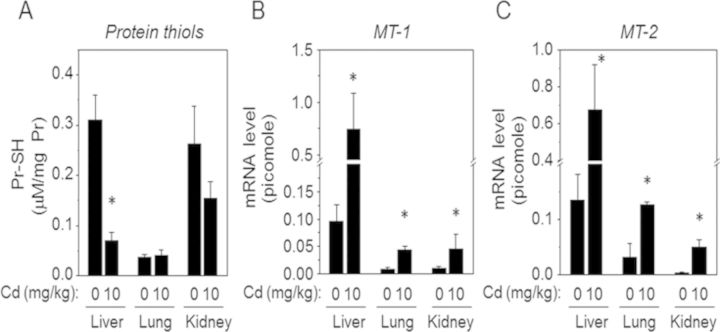
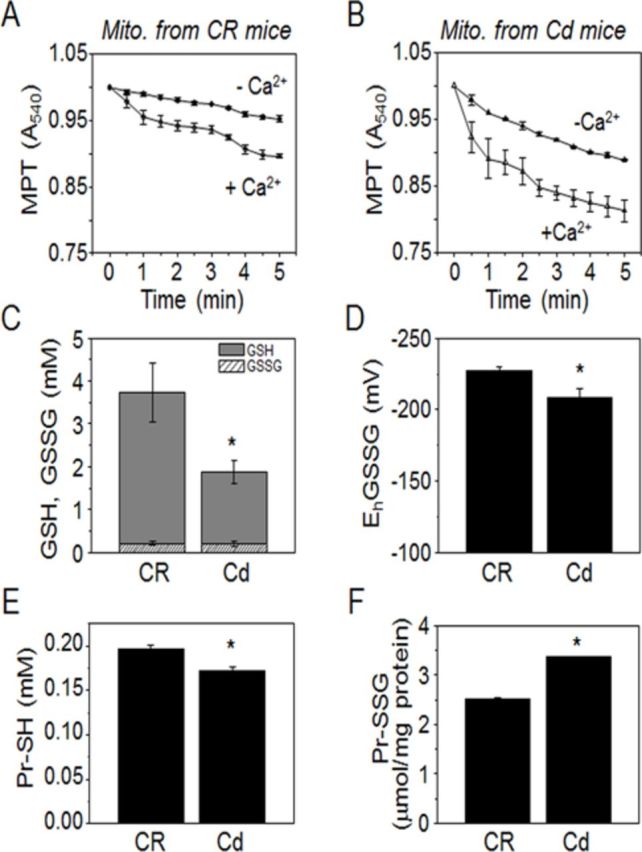
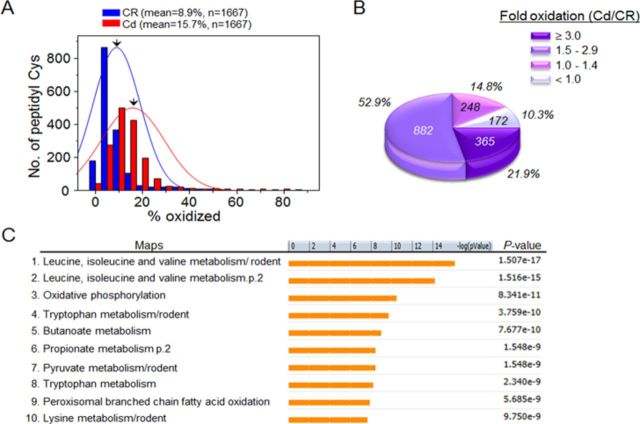
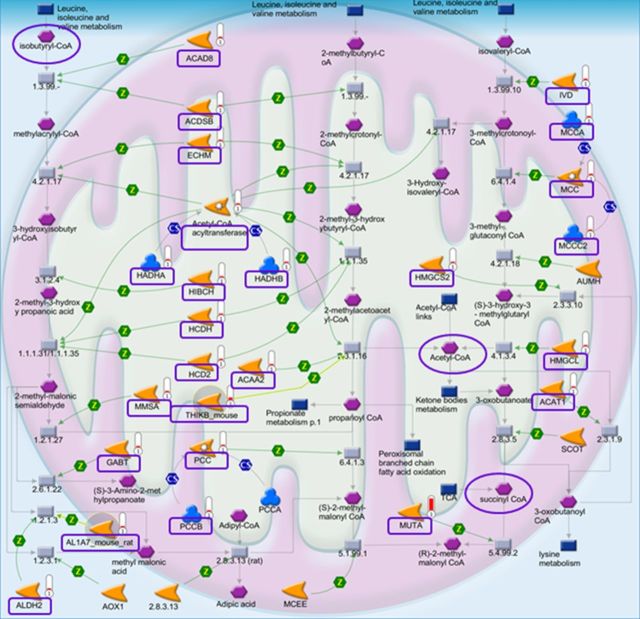
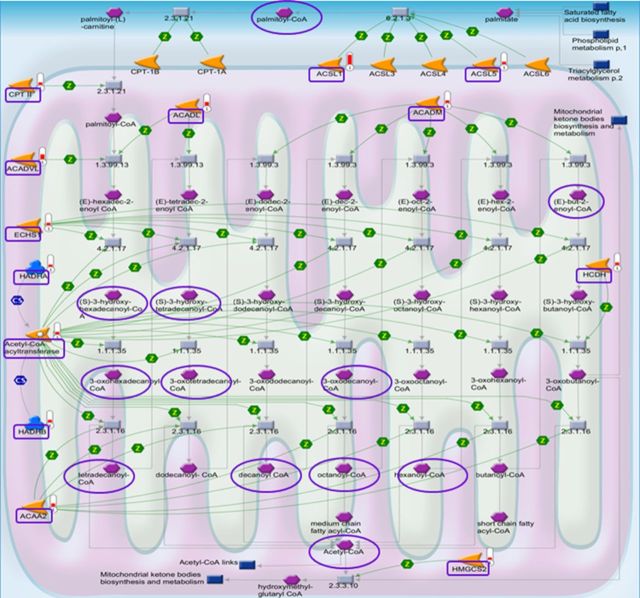
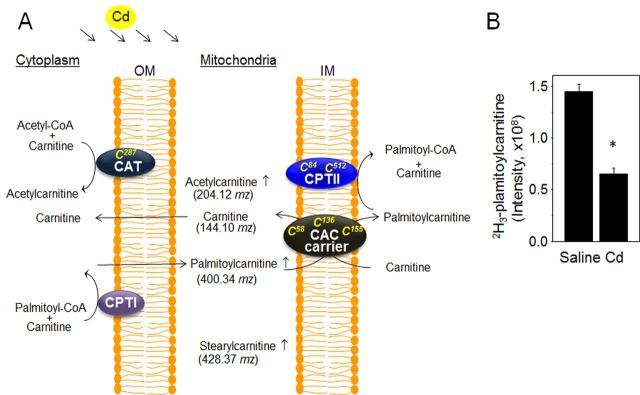
Cadmium (Cd) exposure contributes to human diseases affecting liver, kidney, lung, and other organ systems, but mechanisms underlying the pleotropic nature of these toxicities are poorly understood. Cd accumulates in humans from dietary, environmental (including cigarette smoke), and occupational sources, and has a twenty-year biologic half-life. Our previous mouse and cell studies showed that environmental low-dose Cd exposure altered protein redox states resulting in stimulation of inflammatory signaling and disruption of the actin cytoskeleton system, suggesting that Cd could impact multiple mechanisms of disease. In the current study, we investigated the effects of acute Cd exposure on the redox proteome and metabolome of mouse liver mitochondria to gain insight into associated toxicological mechanisms and functions. We analyzed redox states of liver mitochondrial proteins by redox proteomics using isotope coded affinity tag (ICAT) combined mass spectrometry. Redox ICAT identified 2687 cysteine-containing peptides (peptidyl Cys) of which 1667 peptidyl Cys (657 proteins) were detected in both control and Cd-exposed samples. Of these, 46% (1247 peptidyl Cys, 547 proteins) were oxidized by Cd more than 1.5-fold relative to controls. Bioinformatics analysis using MetaCore software showed that Cd affected 86 pathways, including 24 Cys in proteins functioning in branched chain amino acid (BCAA) and 14 Cys in proteins functioning in fatty acid (acylcarnitine/carnitine) metabolism. Consistently, high-resolution metabolomics data showed that Cd treatment altered levels of BCAA and carnitine metabolites. Together, these results show that mitochondrial protein redox and metabolites are targets in Cd-induced hepatotoxicity. The results further indicate that redox proteomics and metabolomics can be used in an integrated systems approach to investigate complex disease mechanisms.
Keywords: cysteine proteome; environmental toxicant; metabolome; pathway maps; thiol/disulfide redox state.
Figures

Similar articles
-
Actin cytoskeleton redox proteome oxidation by cadmium.Am J Physiol Lung Cell Mol Physiol. 2013 Dec;305(11):L831-43. doi: 10.1152/ajplung.00203.2013. Epub 2013 Sep 27. Am J Physiol Lung Cell Mol Physiol. 2013. PMID: 24077948 Free PMC article.
-
Low-dose cadmium disrupts mitochondrial citric acid cycle and lipid metabolism in mouse lung.Free Radic Biol Med. 2019 Feb 1;131:209-217. doi: 10.1016/j.freeradbiomed.2018.12.005. Epub 2018 Dec 6. Free Radic Biol Med. 2019. PMID: 30529385 Free PMC article.
-
Inhibition of glutathione biosynthesis alters compartmental redox status and the thiol proteome in organogenesis-stage rat conceptuses.Free Radic Biol Med. 2013 Oct;63:325-37. doi: 10.1016/j.freeradbiomed.2013.05.040. Epub 2013 Jun 2. Free Radic Biol Med. 2013. PMID: 23736079 Free PMC article.
-
Proteomic Characterization of Reversible Thiol Oxidations in Proteomes and Proteins.Antioxid Redox Signal. 2017 Mar 1;26(7):329-344. doi: 10.1089/ars.2016.6720. Epub 2016 May 20. Antioxid Redox Signal. 2017. PMID: 27089838 Review.
-
Thiol/disulfide redox states in signaling and sensing.Crit Rev Biochem Mol Biol. 2013 Mar-Apr;48(2):173-81. doi: 10.3109/10409238.2013.764840. Epub 2013 Jan 29. Crit Rev Biochem Mol Biol. 2013. PMID: 23356510 Free PMC article. Review.
Cited by
-
Redox organization of living systems.Free Radic Biol Med. 2024 May 1;217:179-189. doi: 10.1016/j.freeradbiomed.2024.03.008. Epub 2024 Mar 14. Free Radic Biol Med. 2024. PMID: 38490457 Free PMC article. Review.
-
Toxicant-mediated redox control of proteostasis in neurodegeneration.Curr Opin Toxicol. 2019 Feb;13:22-34. doi: 10.1016/j.cotox.2018.12.007. Epub 2018 Dec 28. Curr Opin Toxicol. 2019. PMID: 31602419 Free PMC article.
-
Redox proteomics of tomato in response to Pseudomonas syringae infection.Hortic Res. 2015 Sep 16;2:15043. doi: 10.1038/hortres.2015.43. eCollection 2015. Hortic Res. 2015. PMID: 26504582 Free PMC article.
-
Understanding the Cross-Talk of Redox Metabolism and Fe-S Cluster Biogenesis in Leishmania Through Systems Biology Approach.Front Cell Infect Microbiol. 2019 Feb 4;9:15. doi: 10.3389/fcimb.2019.00015. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 30778378 Free PMC article.
-
Reactive Metabolite-induced Protein Glutathionylation: A Potentially Novel Mechanism Underlying Acetaminophen Hepatotoxicity.Mol Cell Proteomics. 2018 Oct;17(10):2034-2050. doi: 10.1074/mcp.RA118.000875. Epub 2018 Jul 13. Mol Cell Proteomics. 2018. PMID: 30006487 Free PMC article.
References
-
- Andresen B. S., Olpin S., Poorthuis B. J., Scholte H. R., Vianey-Saban C., Wanders R., Ijlst L., Morris A., Pourfarzam M., Bartlett K., et al. Clear correlation of genotype with disease phenotype in very-long-chain acyl-CoA dehydrogenase deficiency. Am. J. Hum. Genet. 1999;64:479–494. - PMC - PubMed
-
- ATSDR. Toxicological profile for cadmium. 2012. Available at: http://www.atsdr.cdc.gov/toxprofiles/tp5.pdf. Accessed February 6, 2006. - PubMed
-
- Bailey N. J., Oven M., Holmes E., Nicholson J. K., Zenk M. H. Metabolomic analysis of the consequences of cadmium exposure in Silene cucubalus cell cultures via 1H NMR spectroscopy and chemometrics. Phytochemistry. 2003;62:851–858. - PubMed
-
- Belyaeva E. A., Korotkov S. M., Saris N. E. In vitro modulation of heavy metal-induced rat liver mitochondria dysfunction: A comparison of copper and mercury with cadmium. J. Trace Elem. Med. Biol. 2011;25(Suppl. 1):S63–S73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
