

Protection from lethal challenge in a neonatal mouse model by circulating recombinant form coxsackievirus A16 vaccine candidates
- PMID: 24496826
- PMCID: PMC3983757
- DOI: 10.1099/vir.0.063560-0
Protection from lethal challenge in a neonatal mouse model by circulating recombinant form coxsackievirus A16 vaccine candidates
Abstract
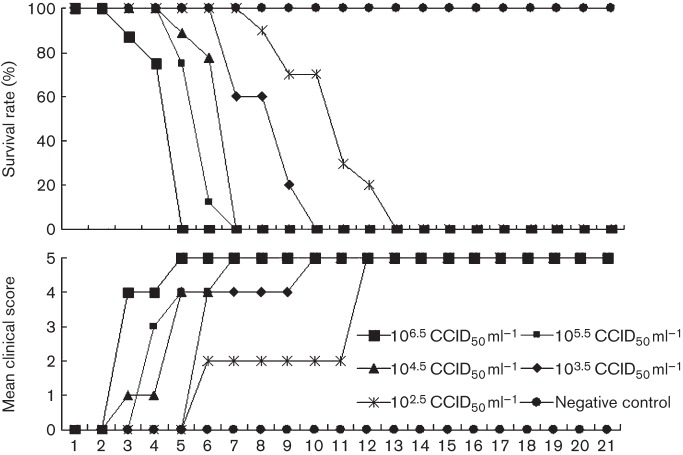
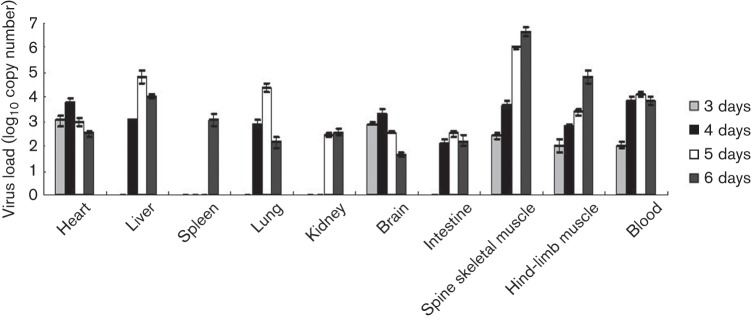
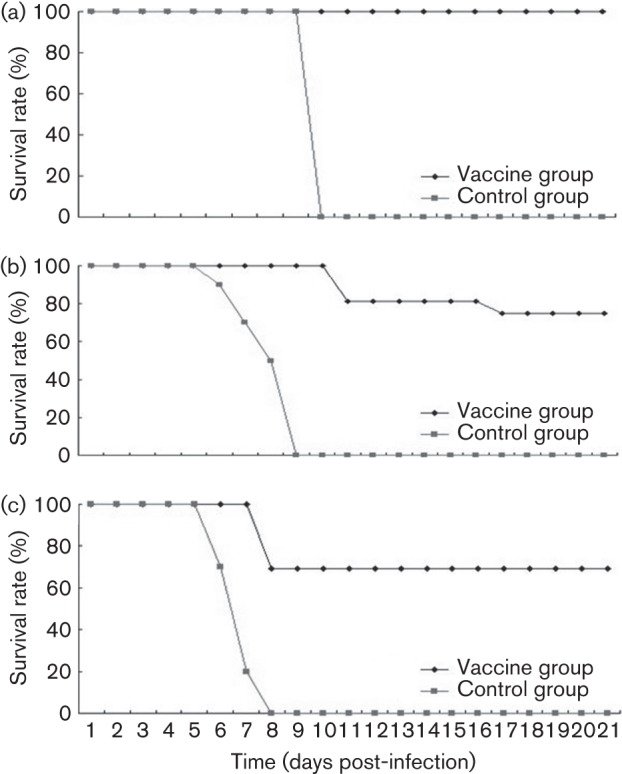
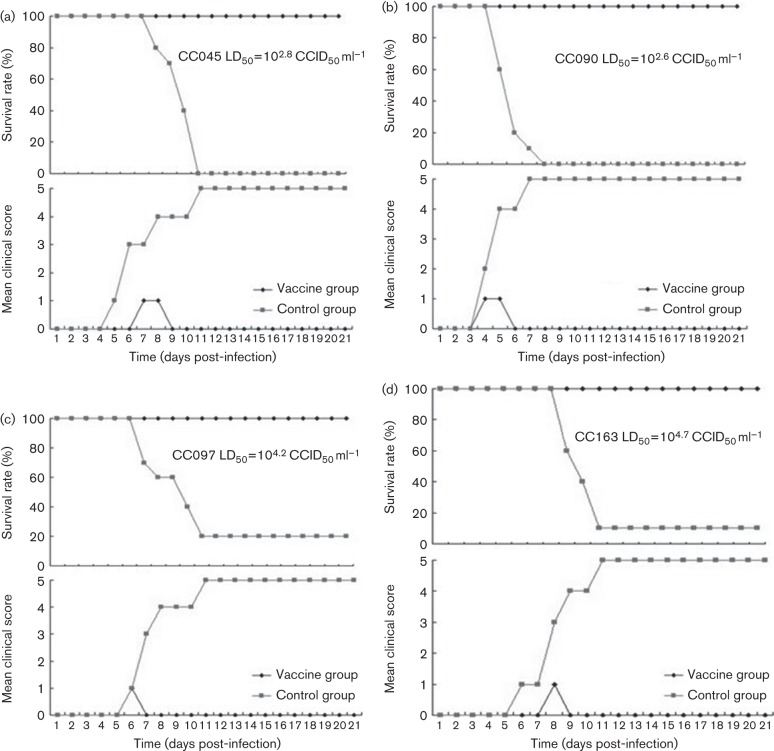
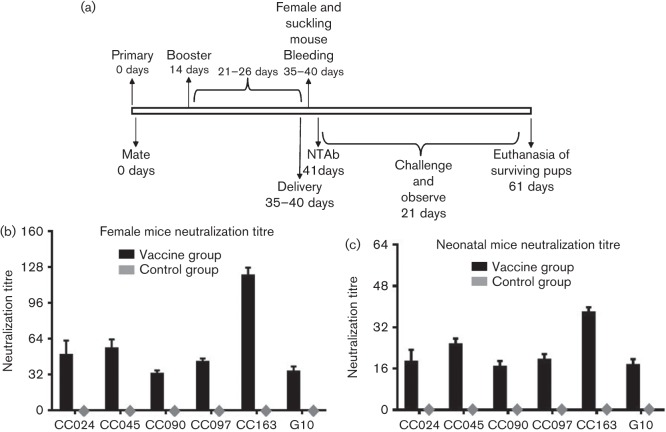
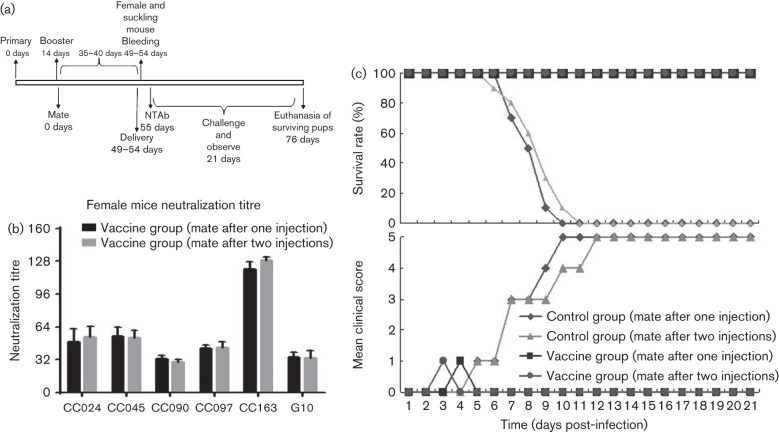
Circulating coxsackievirus A16 (CA16) is a major cause of hand, foot and mouth disease (HFMD) in South-east Asia. At present, there is no vaccine against CA16. Pathogenic animal models that are sensitive to diverse circulating CA16 viruses would be desirable for vaccine development and evaluation. In this study, we isolated and characterized several circulating CA16 viruses from recent HFMD patients. These CA16 viruses currently circulating in humans were highly pathogenic in a newly developed neonatal mouse model; we also observed and analysed the pathogenesis of representative circulating recombinant form CA16 viruses. An inactivated CA16 vaccine candidate, formulated with alum adjuvant and containing submicrogram quantities of viral proteins, protected neonatal mice born to immunized female mice from lethal-dose challenge with a series of CA16 viruses. Further analysis of humoral immunity showed that antibody elicited from both the immunized dams and their pups could neutralize various lethal viruses by a cytopathic effect in vitro. Moreover, viral titres and loads in the tissues of challenged pups in the vaccine group were far lower than those in the control group, and some were undetectable. This lethal-challenge model using pathogenic CA16 viruses and the vaccine candidates that mediated protection in this model could be useful tools for the future development and evaluation of CA16 vaccines.
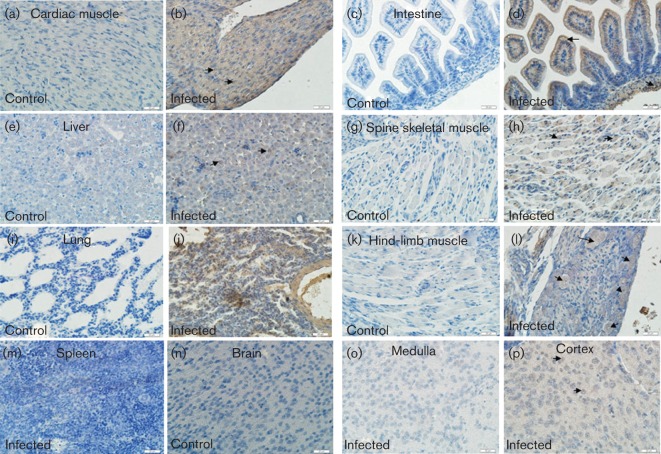
Figures
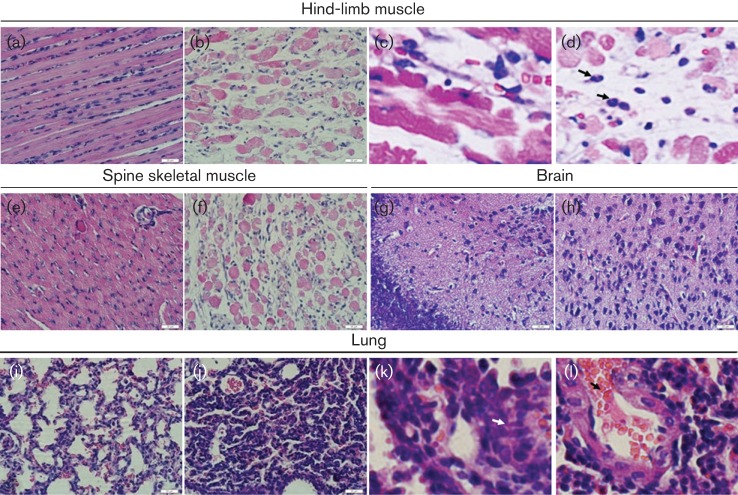
References
-
- Ang L. W., Koh B. K., Chan K. P., Chua L. T., James L., Goh K. T. (2009). Epidemiology and control of hand, foot and mouth disease in Singapore, 2001–2007. Ann Acad Med Singapore 38, 106–112 - PubMed
-
- Bendig J. W., Fleming D. M. (1996). Epidemiological, virological, and clinical features of an epidemic of hand, foot, and mouth disease in England and Wales. Commun Dis Rep CDR Rev 6, R81–R86 - PubMed
-
- Chen S. C., Chang H. L., Yan T. R., Cheng Y. T., Chen K. T. (2007). An eight-year study of epidemiologic features of enterovirus 71 infection in Taiwan. Am J Trop Med Hyg 77, 188–191 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources