

Transcriptional activation of hypoxia-inducible factor-1 (HIF-1) in myeloid cells promotes angiogenesis through VEGF and S100A8
- PMID: 24497508
- PMCID: PMC3932909
- DOI: 10.1073/pnas.1320243111
Transcriptional activation of hypoxia-inducible factor-1 (HIF-1) in myeloid cells promotes angiogenesis through VEGF and S100A8
Abstract
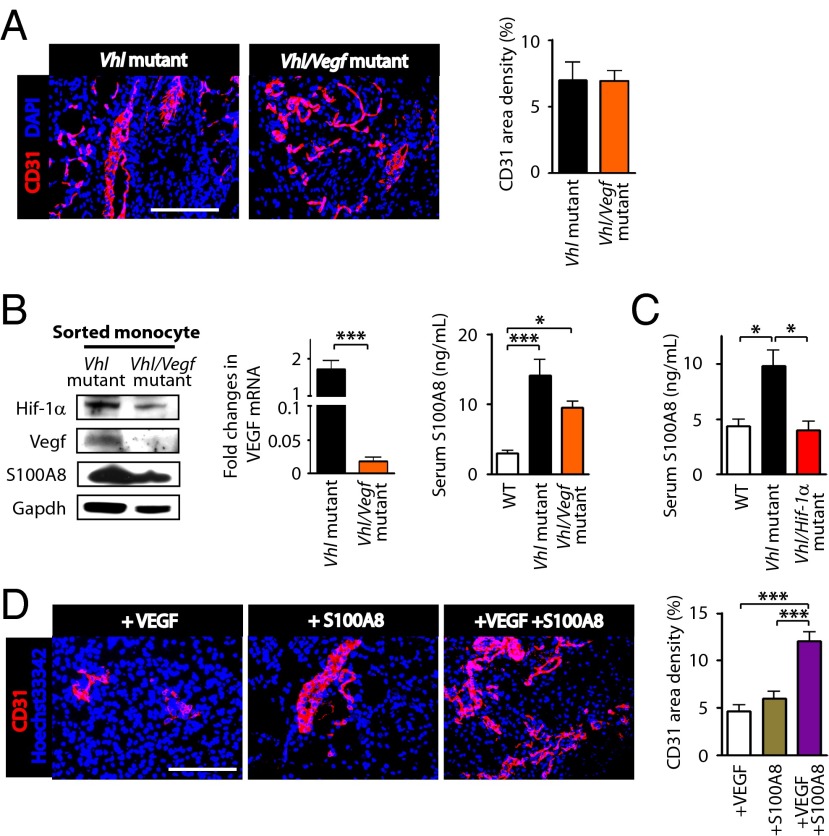
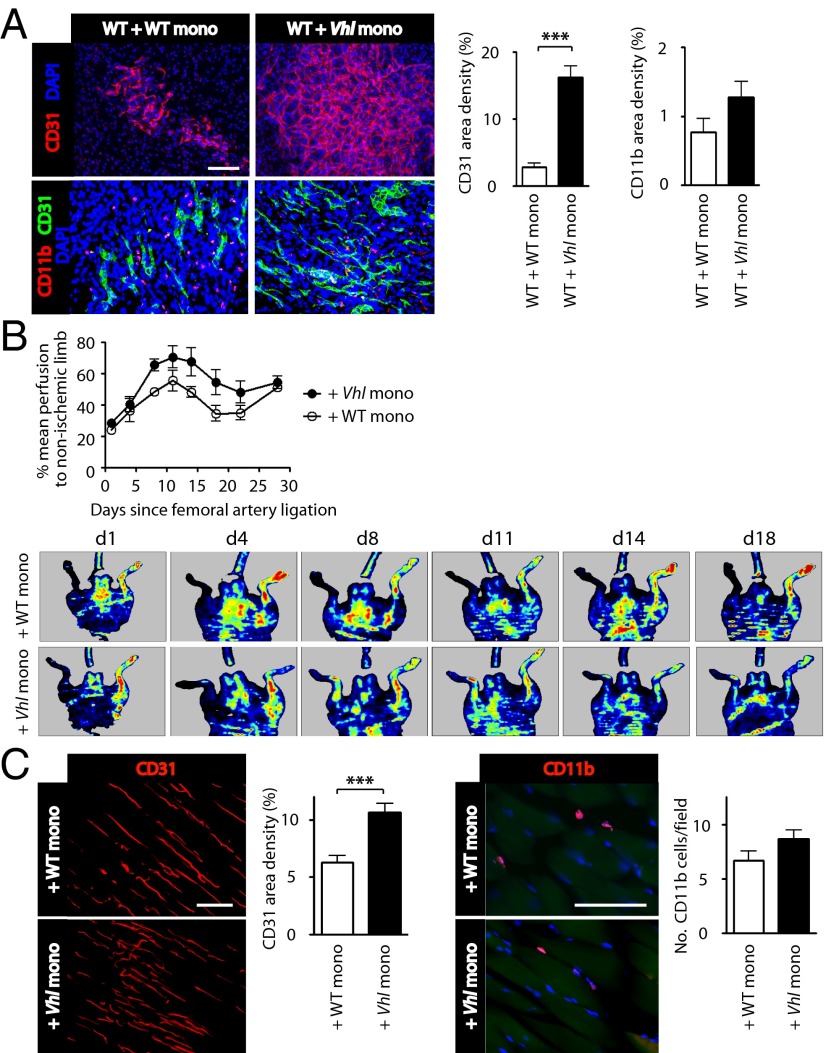
Emerging evidence indicates that myeloid cells are essential for promoting new blood vessel formation by secreting various angiogenic factors. Given that hypoxia-inducible factor (HIF) is a critical regulator for angiogenesis, we questioned whether HIF in myeloid cells also plays a role in promoting angiogenesis. To address this question, we generated a unique strain of myeloid-specific knockout mice targeting HIF pathways using human S100A8 as a myeloid-specific promoter. We observed that mutant mice where HIF-1 is transcriptionally activated in myeloid cells (by deletion of the von Hippel-Lindau gene) resulted in erythema, enhanced neovascularization in matrigel plugs, and increased production of vascular endothelial growth factor (VEGF) in the bone marrow, all of which were completely abrogated by either genetic or pharmacological inactivation of HIF-1. We further found that monocytes were the major effector producing VEGF and S100A8 proteins driving neovascularization in matrigel. Moreover, by using a mouse model of hindlimb ischemia we observed significantly improved blood flow in mice intramuscularly injected with HIF-1-activated monocytes. This study therefore demonstrates that HIF-1 activation in myeloid cells promotes angiogenesis through VEGF and S100A8 and that this may become an attractive therapeutic strategy to treat diseases with vascular defects.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
