

A role for mitochondrial phosphoenolpyruvate carboxykinase (PEPCK-M) in the regulation of hepatic gluconeogenesis
- PMID: 24497630
- PMCID: PMC3953244
- DOI: 10.1074/jbc.C113.544759
A role for mitochondrial phosphoenolpyruvate carboxykinase (PEPCK-M) in the regulation of hepatic gluconeogenesis
Abstract
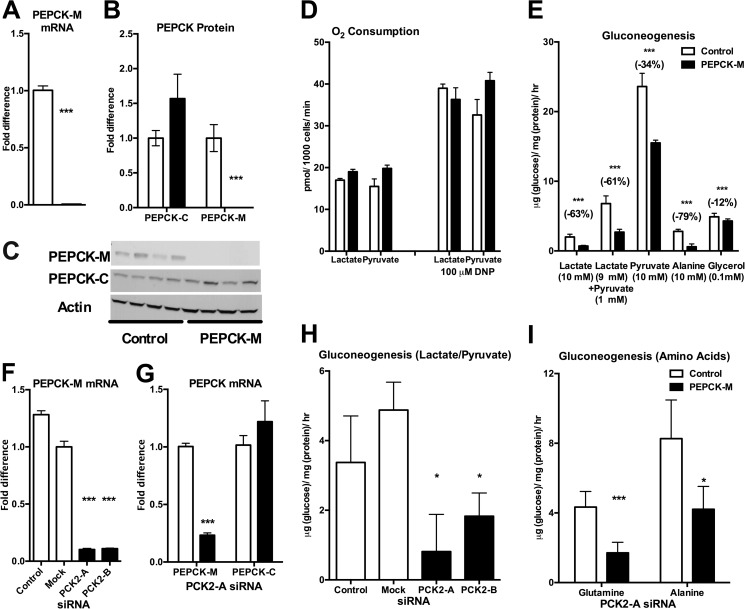
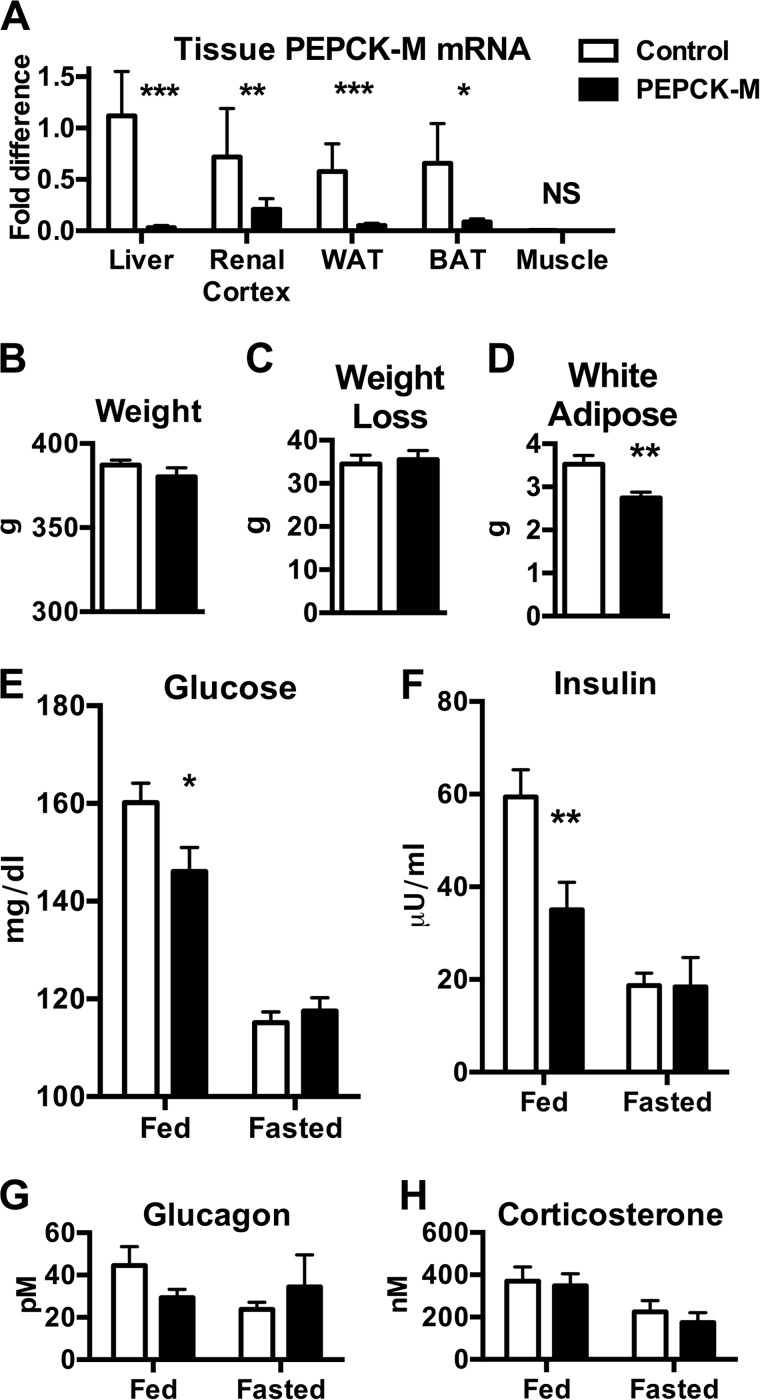
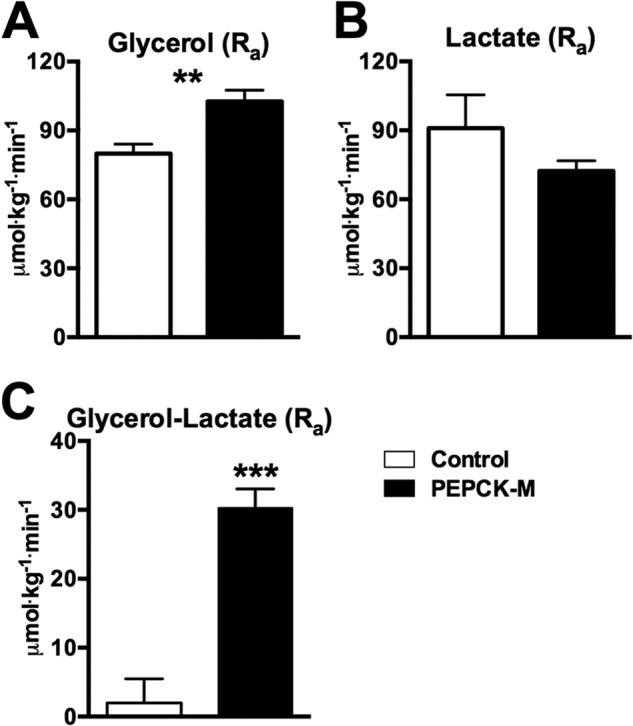
Synthesis of phosphoenolpyruvate (PEP) from oxaloacetate is an absolute requirement for gluconeogenesis from mitochondrial substrates. Generally, this reaction has solely been attributed to the cytosolic isoform of PEPCK (PEPCK-C), although loss of the mitochondrial isoform (PEPCK-M) has never been assessed. Despite catalyzing the same reaction, to date the only significant role reported in mammals for the mitochondrial isoform is as a glucose sensor necessary for insulin secretion. We hypothesized that this nutrient-sensing mitochondrial GTP-dependent pathway contributes importantly to gluconeogenesis. PEPCK-M was acutely silenced in gluconeogenic tissues of rats using antisense oligonucleotides both in vivo and in isolated hepatocytes. Silencing PEPCK-M lowers plasma glucose, insulin, and triglycerides, reduces white adipose, and depletes hepatic glycogen, but raises lactate. There is a switch of gluconeogenic substrate preference to glycerol that quantitatively accounts for a third of glucose production. In contrast to the severe mitochondrial deficiency characteristic of PEPCK-C knock-out livers, hepatocytes from PEPCK-M-deficient livers maintained normal oxidative function. Consistent with its predicted role, gluconeogenesis rates from hepatocytes lacking PEPCK-M are severely reduced for lactate, alanine, and glutamine, but not for pyruvate and glycerol. Thus, PEPCK-M has a direct role in fasted and fed glucose homeostasis, and this mitochondrial GTP-dependent pathway should be reconsidered for its involvement in both normal and diabetic metabolism.
Keywords: Diabetes; GTPase; Gluconeogenesis; Glyceroneogenesis; Intermediary Metabolism; Metabolic Regulation; Metabolic Tracers; Metabolism; Mitochondrial Metabolism; Phosphoenolpyruvate Carboxykinase.
Figures
References
-
- Gastaldelli A., Miyazaki Y., Pettiti M., Buzzigoli E., Mahankali S., Ferrannini E., DeFronzo R. A. (2004) Separate contribution of diabetes, total fat mass, and fat topography to glucose production, gluconeogenesis, and glycogenolysis. J. Clin. Endocrinol. Metab. 89, 3914–3921 - PubMed
-
- Maggs D. G., Buchanan T. A., Burant C. F., Cline G., Gumbiner B., Hsueh W. A., Inzucchi S., Kelley D., Nolan J., Olefsky J. M., Polonsky K. S., Silver D., Valiquett T. R., Shulman G. I. (1998) Metabolic effects of troglitazone monotherapy in type 2 diabetes mellitus. A randomized, double-blind, placebo-controlled trial. Ann. Intern. Med. 128, 176–185 - PubMed
-
- Owen O. E., Kalhan S. C., Hanson R. W. (2002) The key role of anaplerosis and cataplerosis for citric acid cycle function. J. Biol. Chem. 277, 30409–30412 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R24 DK-085638/DK/NIDDK NIH HHS/United States
- R01 DK-40936/DK/NIDDK NIH HHS/United States
- P30 DK079310/DK/NIDDK NIH HHS/United States
- P30 DK45735/DK/NIDDK NIH HHS/United States
- U24 DK059635/DK/NIDDK NIH HHS/United States
- R01 DK040936/DK/NIDDK NIH HHS/United States
- R24 DK085638/DK/NIDDK NIH HHS/United States
- P30 DK79310-01/DK/NIDDK NIH HHS/United States
- U24 DK-059635/DK/NIDDK NIH HHS/United States
- P30 DK034989/DK/NIDDK NIH HHS/United States
- K08 DK080142/DK/NIDDK NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P30 DK34989/DK/NIDDK NIH HHS/United States
- P30 DK045735/DK/NIDDK NIH HHS/United States
- R01 DK092606/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
