

β-hairpin-mediated formation of structurally distinct multimers of neurotoxic prion peptides
- PMID: 24498083
- PMCID: PMC3909104
- DOI: 10.1371/journal.pone.0087354
β-hairpin-mediated formation of structurally distinct multimers of neurotoxic prion peptides
Abstract
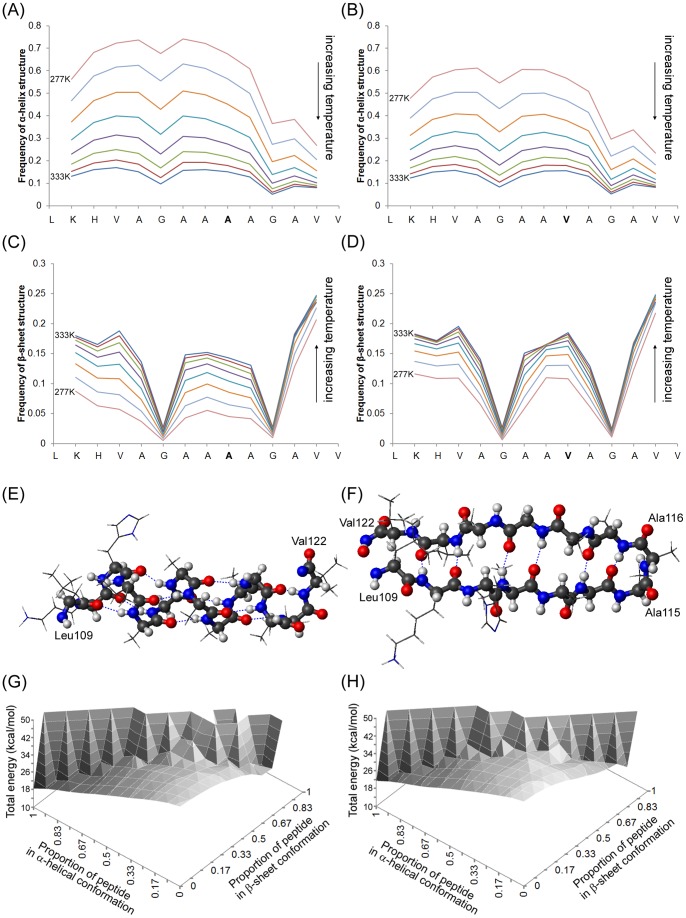
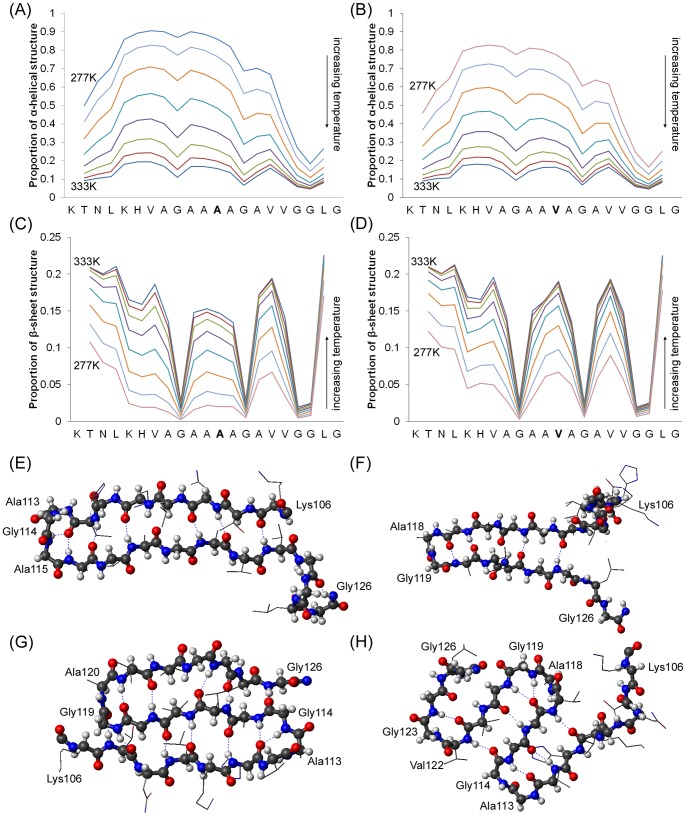
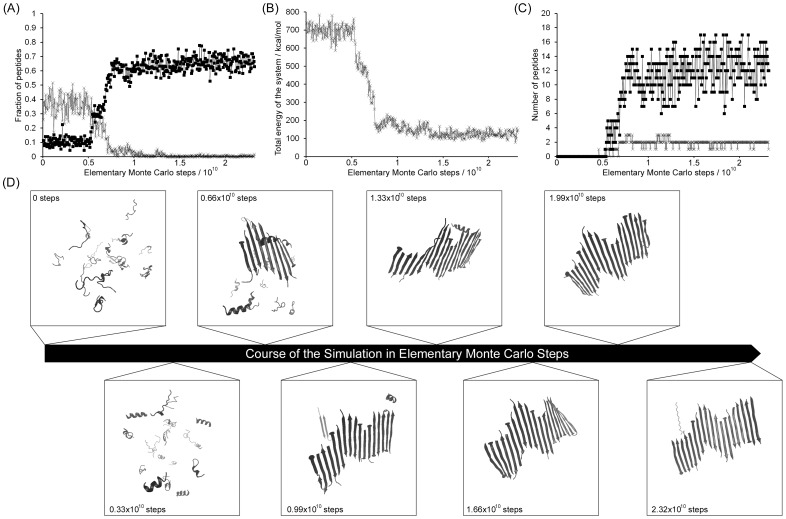
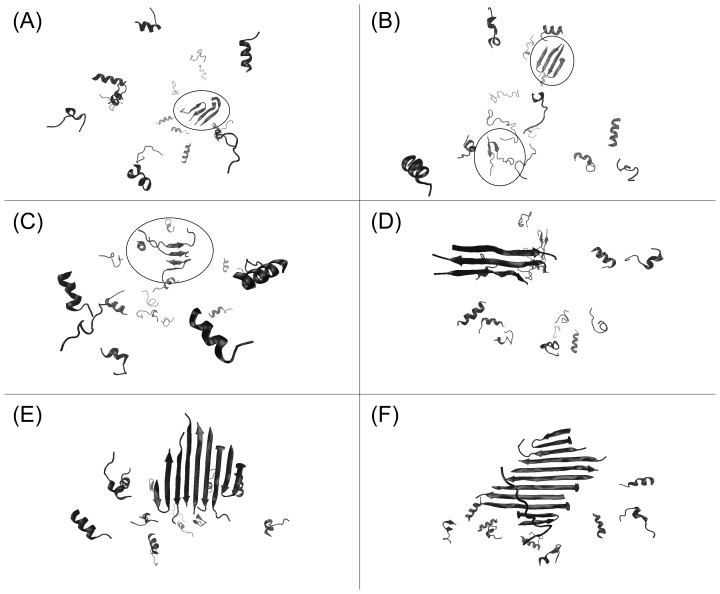
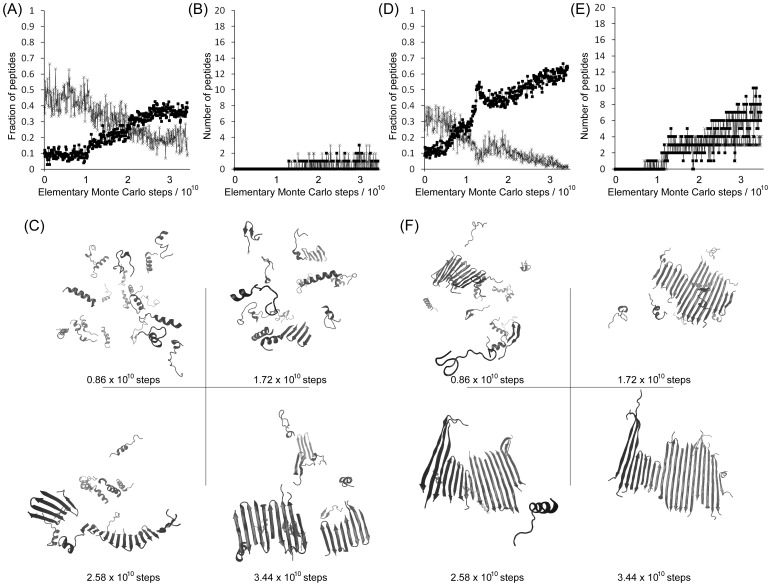
Protein misfolding disorders are associated with conformational changes in specific proteins, leading to the formation of potentially neurotoxic amyloid fibrils. During pathogenesis of prion disease, the prion protein misfolds into β-sheet rich, protease-resistant isoforms. A key, hydrophobic domain within the prion protein, comprising residues 109-122, recapitulates many properties of the full protein, such as helix-to-sheet structural transition, formation of fibrils and cytotoxicity of the misfolded isoform. Using all-atom, molecular simulations, it is demonstrated that the monomeric 109-122 peptide has a preference for α-helical conformations, but that this peptide can also form β-hairpin structures resulting from turns around specific glycine residues of the peptide. Altering a single amino acid within the 109-122 peptide (A117V, associated with familial prion disease) increases the prevalence of β-hairpin formation and these observations are replicated in a longer peptide, comprising residues 106-126. Multi-molecule simulations of aggregation yield different assemblies of peptide molecules composed of conformationally-distinct monomer units. Small molecular assemblies, consistent with oligomers, comprise peptide monomers in a β-hairpin-like conformation and in many simulations appear to exist only transiently. Conversely, larger assemblies are comprised of extended peptides in predominately antiparallel β-sheets and are stable relative to the length of the simulations. These larger assemblies are consistent with amyloid fibrils, show cross-β structure and can form through elongation of monomer units within pre-existing oligomers. In some simulations, assemblies containing both β-hairpin and linear peptides are evident. Thus, in this work oligomers are on pathway to fibril formation and a preference for β-hairpin structure should enhance oligomer formation whilst inhibiting maturation into fibrils. These simulations provide an important new atomic-level model for the formation of oligomers and fibrils of the prion protein and suggest that stabilization of β-hairpin structure may enhance cellular toxicity by altering the balance between oligomeric and fibrillar protein assemblies.
Conflict of interest statement
Figures
References
-
- Chiti F, Dobson CM (2006) Protein misfolding, functional amyloid, and human disease. Annual Review of Biochemistry 75: 333–366. - PubMed
-
- Clark A, Dekoning EJP, Hattersley AT, Hansen BC, Yajnik CS, et al. (1995) Pancreatic Pathology in Non-Insulin-Dependent Diabetes (Niddm). Diabetes Research and Clinical Practice 28: S39–S47. - PubMed
-
- Yu LP, Edalji R, Harlan JE, Holzman TF, Lopez AP, et al. (2009) Structural Characterization of a Soluble Amyloid beta-Peptide Oligomer. Biochemistry 48: 1870–1877. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
