

TAF4 inactivation reveals the 3 dimensional growth promoting activities of collagen 6A3
- PMID: 24498316
- PMCID: PMC3911972
- DOI: 10.1371/journal.pone.0087365
TAF4 inactivation reveals the 3 dimensional growth promoting activities of collagen 6A3
Abstract
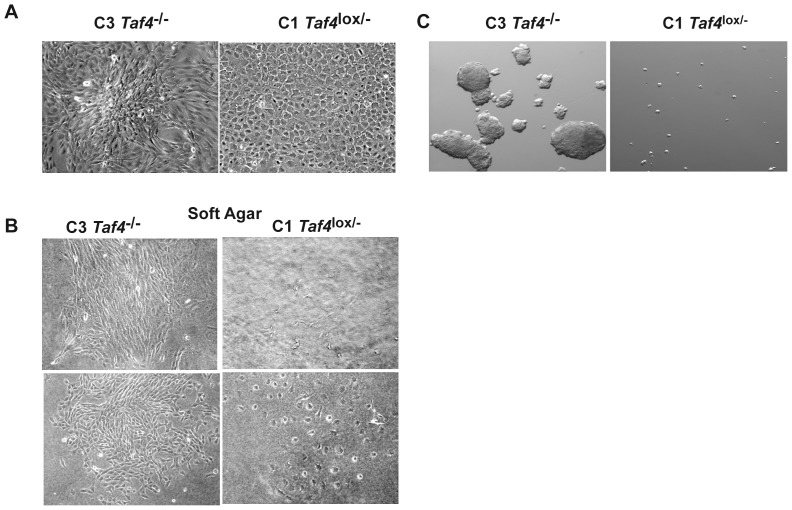
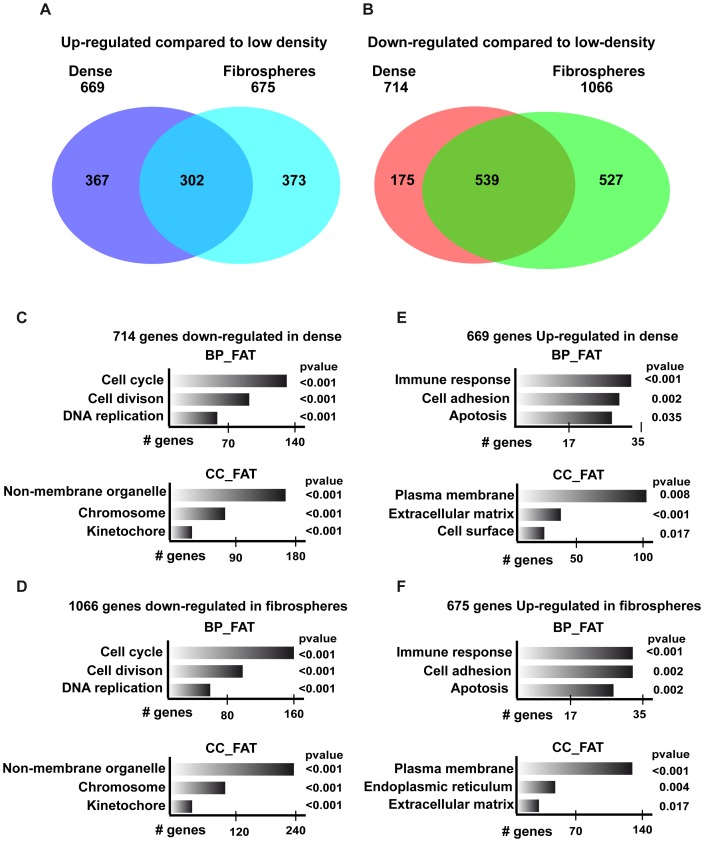
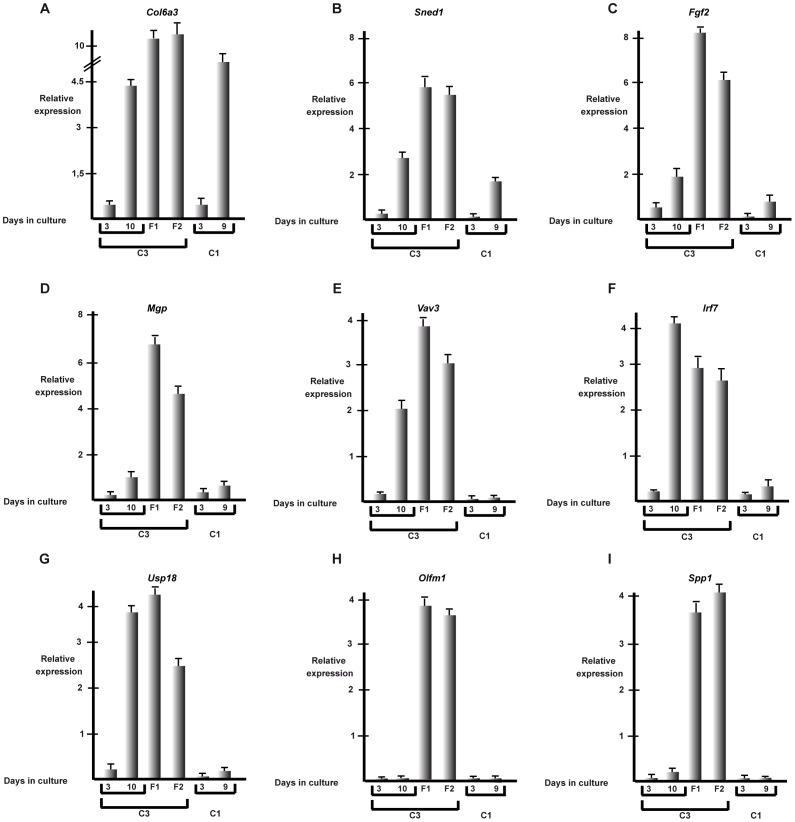
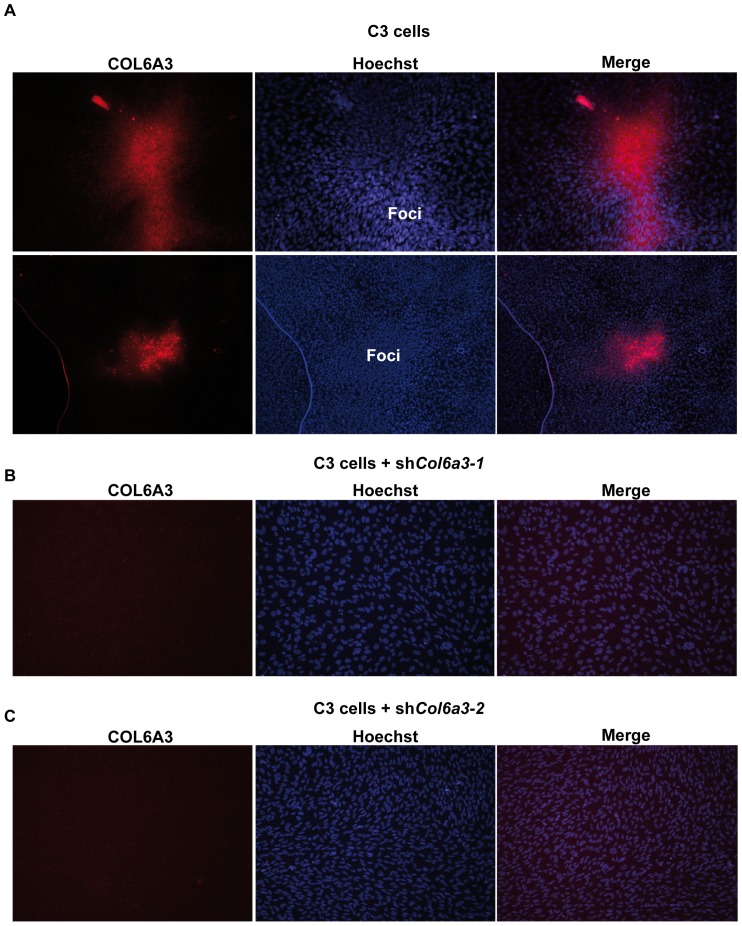
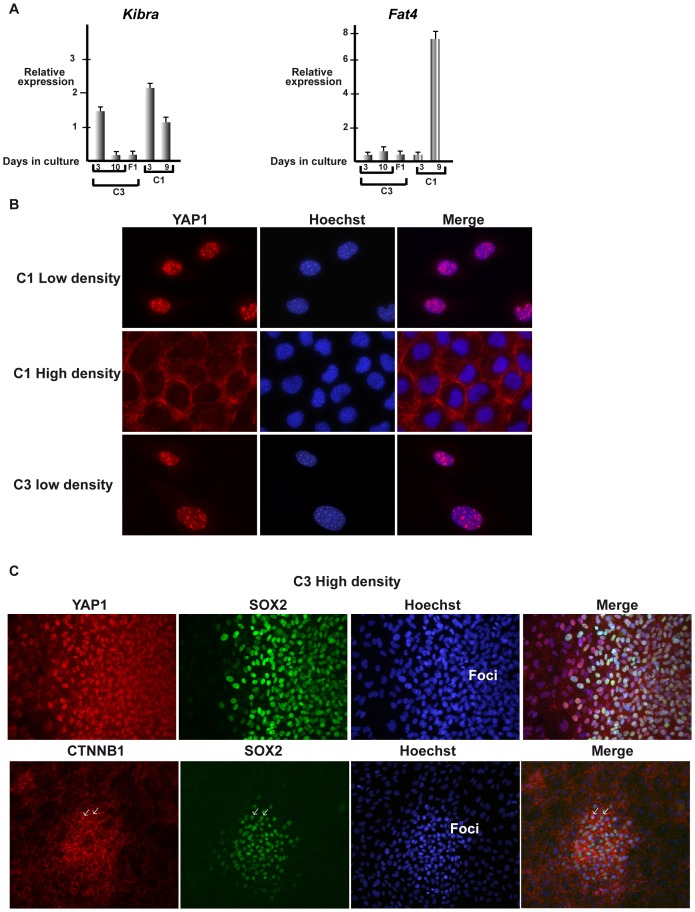
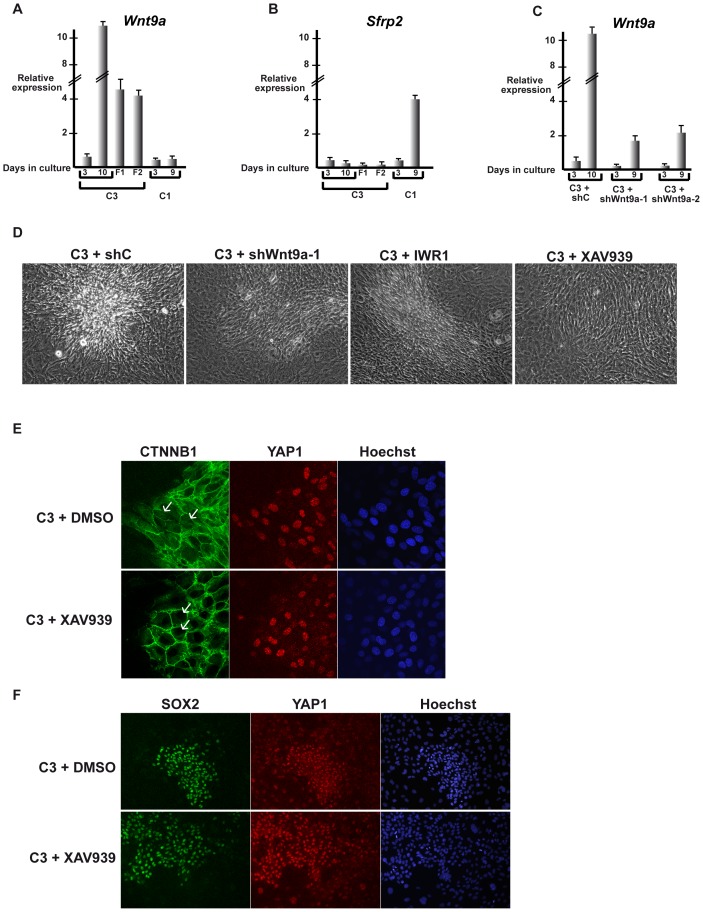
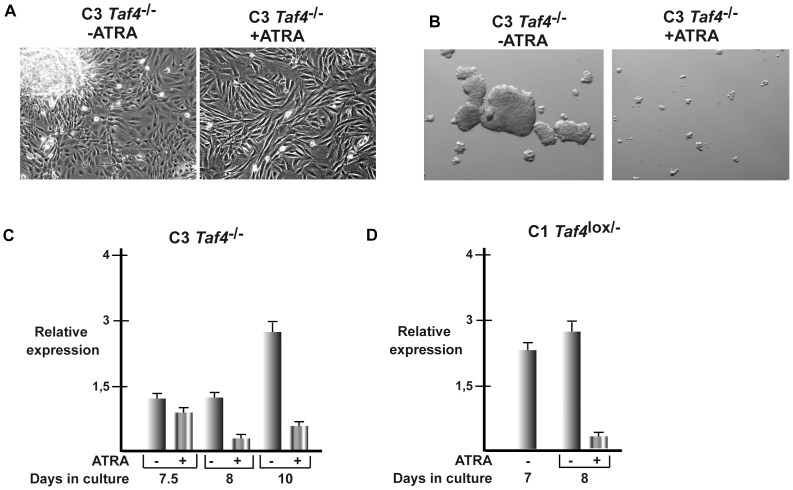
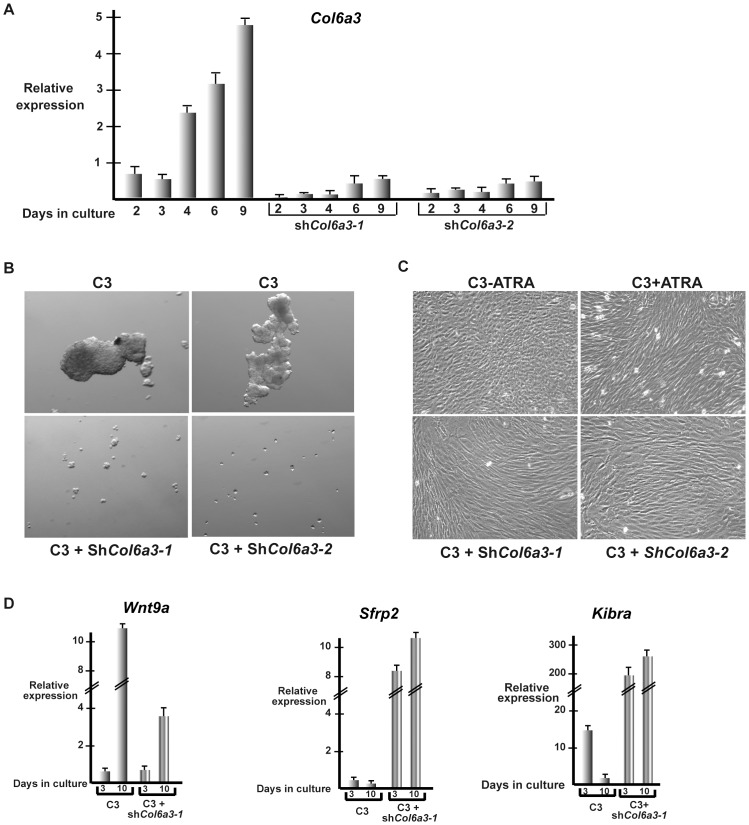
Collagen 6A3 (Col6a3), a component of extracellular matrix, is often up-regulated in tumours and is believed to play a pro-oncogenic role. However the mechanisms of its tumorigenic activity are poorly understood. We show here that Col6a3 is highly expressed in densely growing mouse embryonic fibroblasts (MEFs). In MEFs where the TAF4 subunit of general transcription factor IID (TFIID) has been inactivated, elevated Col6a3 expression prevents contact inhibition promoting their 3 dimensional growth as foci and fibrospheres. Analyses of gene expression in densely growing Taf4(-/-) MEFs revealed repression of the Hippo pathway and activation of Wnt signalling. The Hippo activator Kibra/Wwc1 is repressed under dense conditions in Taf4(-/-) MEFs, leading to nuclear accumulation of the proliferation factor YAP1 in the cells forming 3D foci. At the same time, Wnt9a is activated and the Sfrp2 antagonist of Wnt signalling is repressed. Surprisingly, treatment of Taf4(-/-) MEFs with all-trans retinoic acid (ATRA) restores contact inhibition suppressing 3D growth. ATRA represses Col6a3 expression independently of TAF4 expression and Col6a3 silencing is sufficient to restore contact inhibition in Taf4(-/-) MEFs and to suppress 3D growth by reactivating Kibra expression to induce Hippo signalling and by inducing Sfrp2 expression to antagonize Wnt signalling. All together, these results reveal a critical role for Col6a3 in regulating both Hippo and Wnt signalling to promote 3D growth, and show that the TFIID subunit TAF4 is essential to restrain the growth promoting properties of Col6a3. Our data provide new insight into the role of extra cellular matrix components in regulating cell growth.
Conflict of interest statement
Figures

References
-
- Freiman RN, Albright SR, Zheng S, Sha WC, Hammer RE, et al. (2001) Requirement of tissue-selective TBP-associated factor TAFII105 in ovarian development. Science 293: 2084–2087. - PubMed
-
- Fadloun A, Kobi D, Pointud JC, Indra AK, Teletin M, et al. (2007) The TFIID subunit TAF4 regulates keratinocyte proliferation and has cell-autonomous and non-cell-autonomous tumour suppressor activity in mouse epidermis. Development 134: 2947–2958. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
