

Plasmodium falciparum Rab5B is an N-terminally myristoylated Rab GTPase that is targeted to the parasite's plasma and food vacuole membranes
- PMID: 24498355
- PMCID: PMC3912013
- DOI: 10.1371/journal.pone.0087695
Plasmodium falciparum Rab5B is an N-terminally myristoylated Rab GTPase that is targeted to the parasite's plasma and food vacuole membranes
Abstract
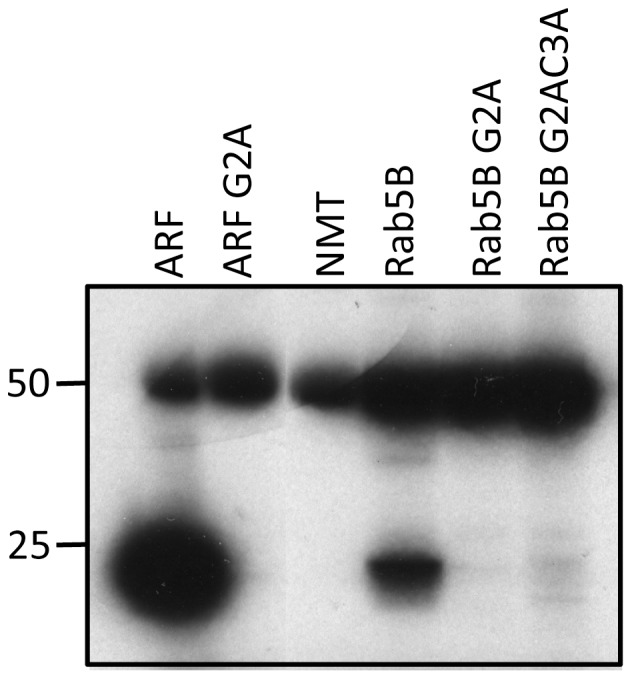
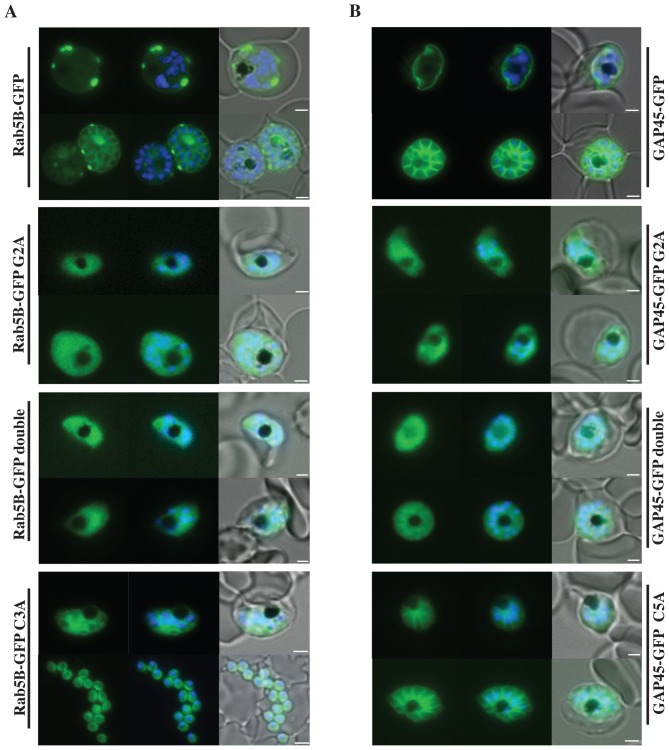
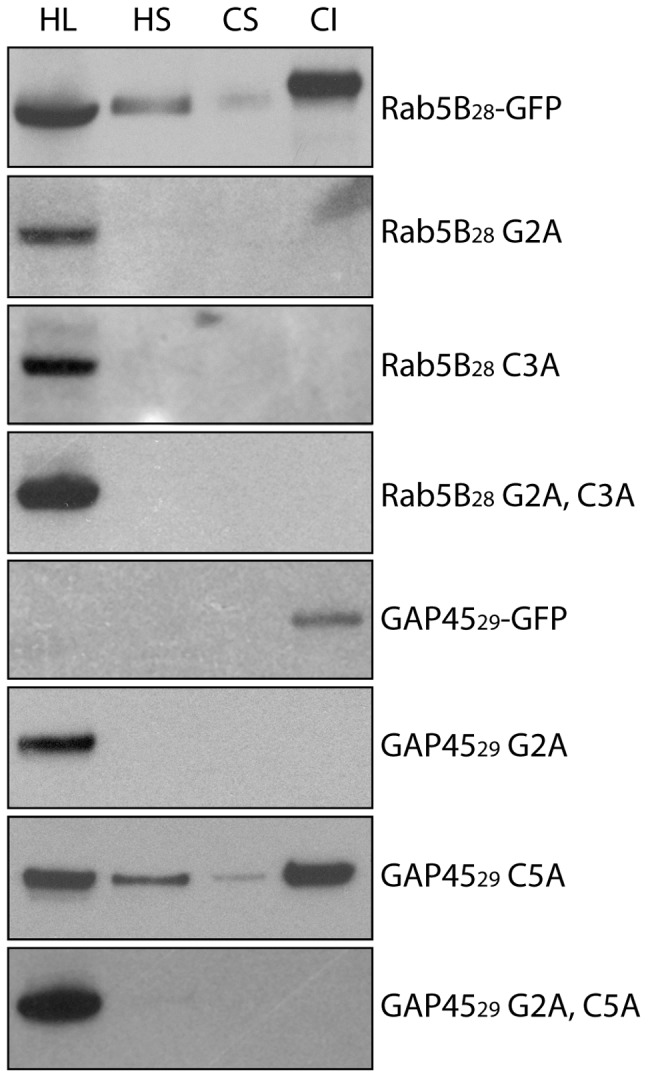
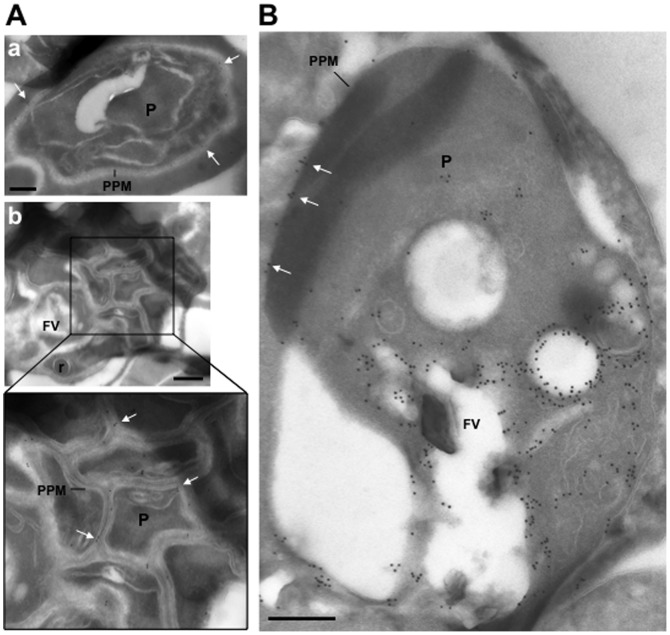
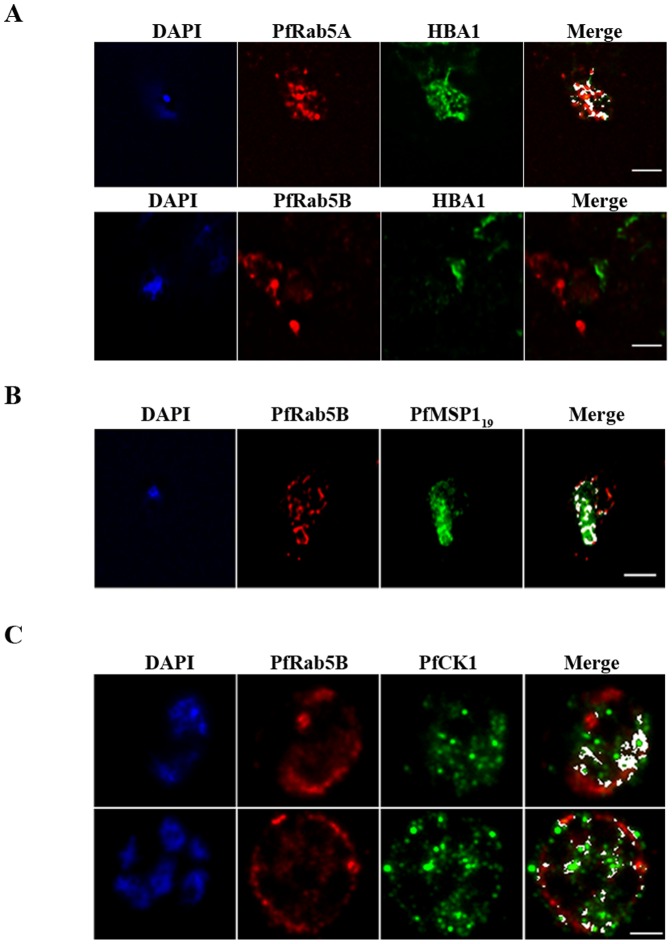
Plasmodium falciparum (Pf) has a family of 11 Rab GTPases to regulate its vesicular transport. However, PfRab5B is unique in lacking a C-terminal geranyl-geranylation motif, while having N-terminal palmitoylation and myristoylation motifs. We show that the N-terminal glycine is required for PfRab5B myristoylation in vitro and when an N-terminal PfRab5B fragment possessing both acylation motifs is fused to GFP and expressed in transgenic P. falciparum parasites, the chimeric PfRab5B protein localizes to the plasma membrane. Upon substitution of the modified glycine by alanine the staining becomes diffuse and GFP is found in soluble subcellular fractions. Immuno-electron microscopy shows endogenous PfRab5B decorating the parasite's plasma and food vacuole membranes. Using reverse genetics rab5b couldn't be deleted from the haploid genome of asexual blood stage P. berghei parasites. The failure of PbRab5A or PbRab5C to complement for loss of PbRab5B function indicates non-overlapping roles for the three Plasmodium Rab5s, with PfRab5B involved in trafficking MSP1 to the food vacuole membrane and CK1 to the plasma membrane. We discuss similarities between Plasmodium Rab5B and Arabidopsis thaliana ARA6, a similarly unusual Rab5-like GTPase of plants.
Conflict of interest statement
Figures
References
-
- White NJ, Pukrittayakamee S, Hien TT, Faiz MA, Mokuolu OA, et al. (2013) Malaria. Lancet - PubMed
-
- de Castro FA, Ward GE, Jambou R, Attal G, Mayau V, et al. (1996) Identification of a family of Rab G-proteins in Plasmodium falciparum and a detailed characterisation of pfrab6. Mol Biochem Parasitol 80: 77–88. - PubMed
-
- Ward GE, Tilney LG, Langsley G (1997) Rab GTPases and the unusual secretory pathway of plasmodium. Parasitol Today 13: 57–62. - PubMed
-
- Zerial M, McBride H (2001) Rab proteins as membrane organizers. Nat Rev Mol Cell Biol 2: 107–117. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
