

Concentration-dependent exchange of replication protein A on single-stranded DNA revealed by single-molecule imaging
- PMID: 24498402
- PMCID: PMC3912175
- DOI: 10.1371/journal.pone.0087922
Concentration-dependent exchange of replication protein A on single-stranded DNA revealed by single-molecule imaging
Abstract
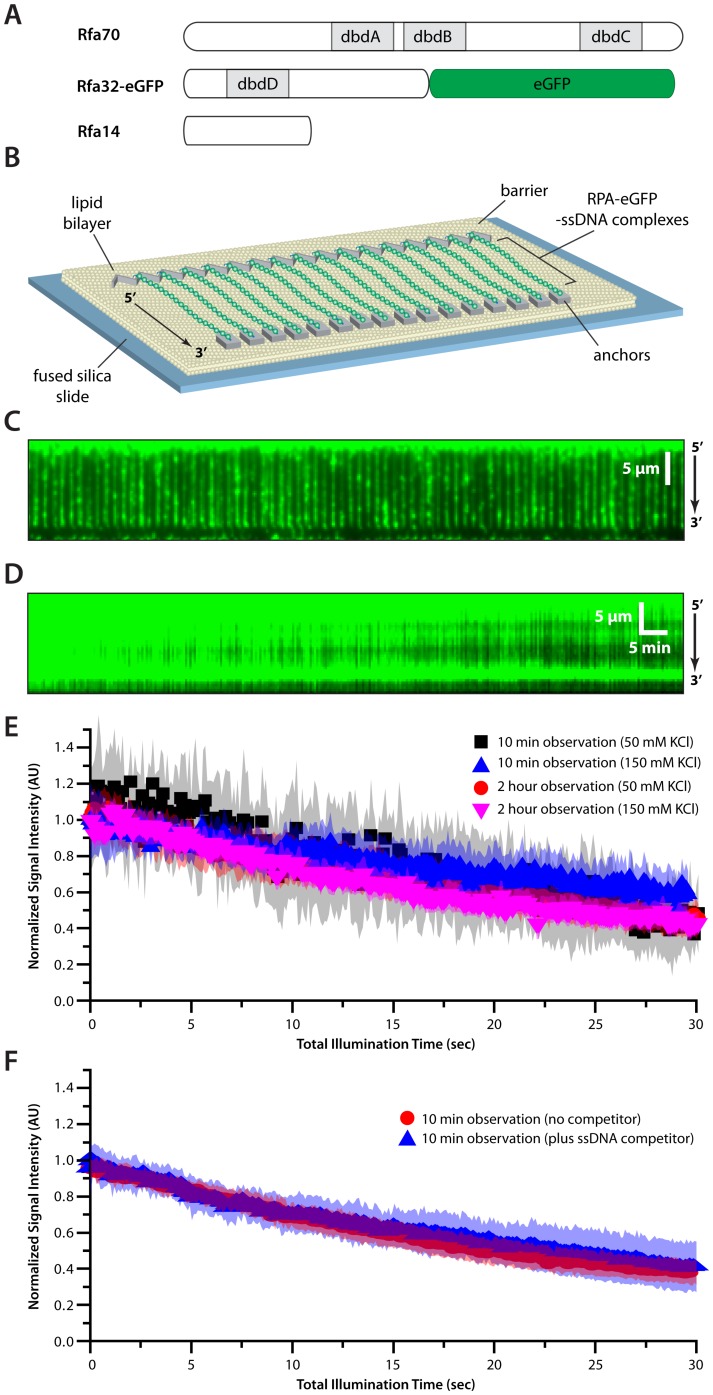
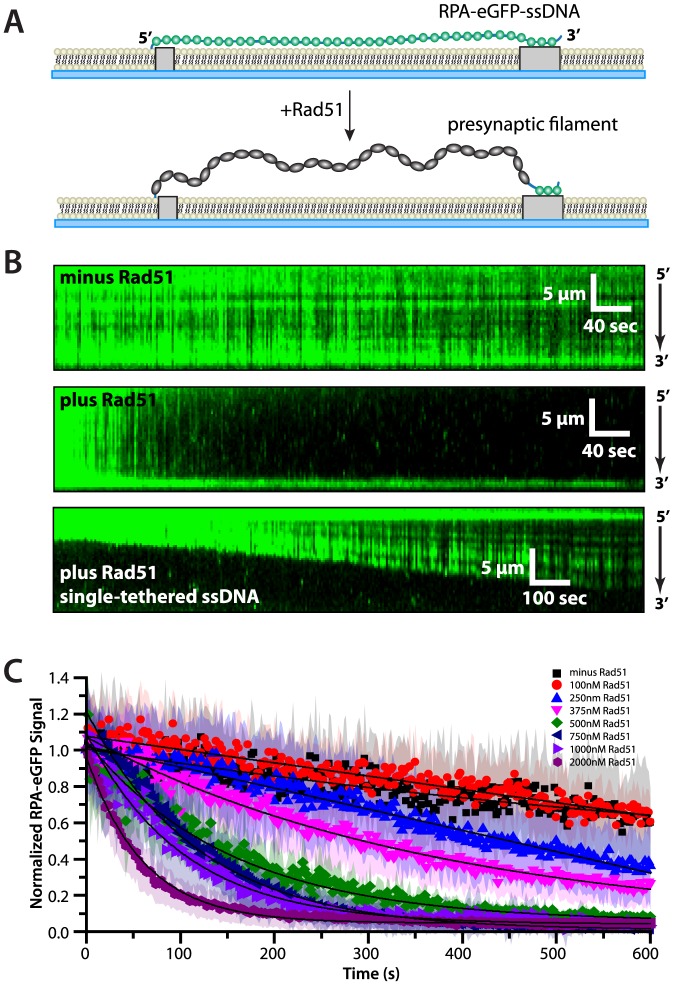
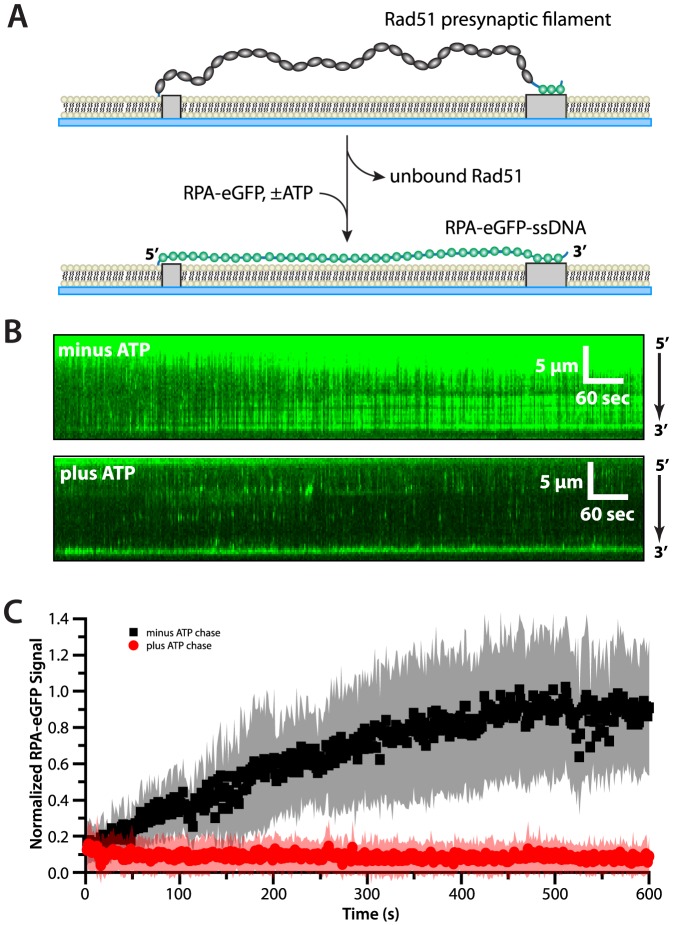
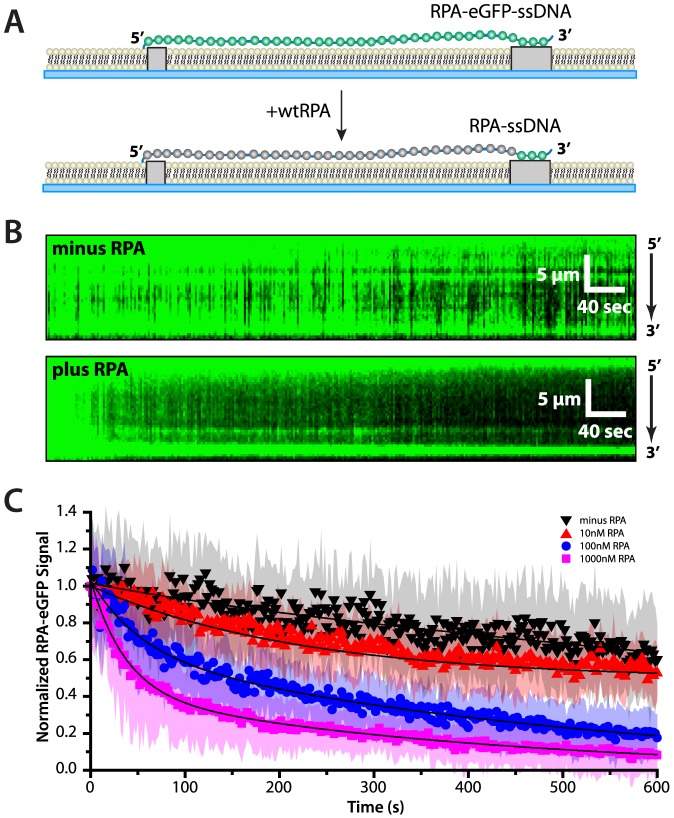
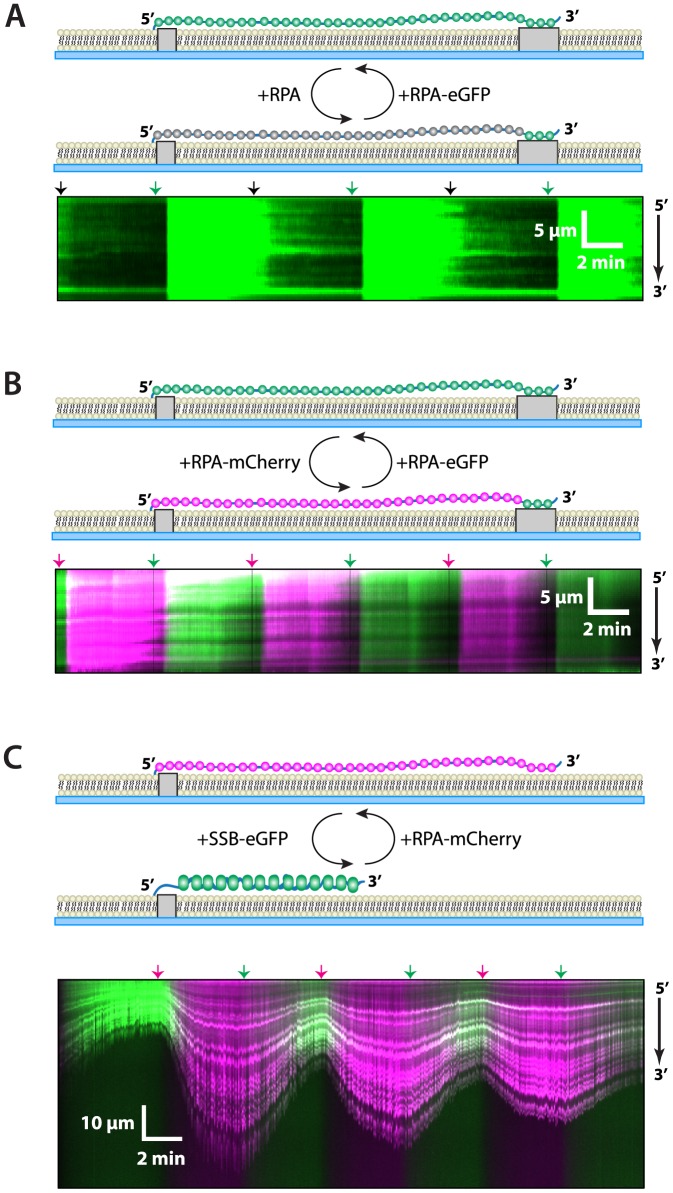
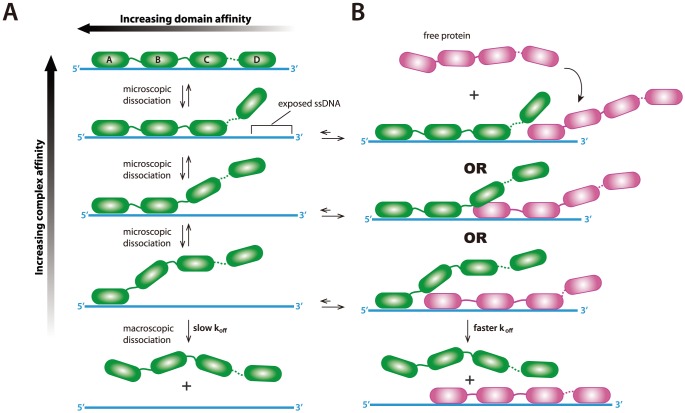
Replication protein A (RPA) is a ubiquitous eukaryotic single-stranded DNA (ssDNA) binding protein necessary for all aspects of DNA metabolism involving an ssDNA intermediate, including DNA replication, repair, recombination, DNA damage response and checkpoint activation, and telomere maintenance. The role of RPA in most of these reactions is to protect the ssDNA until it can be delivered to downstream enzymes. Therefore a crucial feature of RPA is that it must bind very tightly to ssDNA, but must also be easily displaced from ssDNA to allow other proteins to gain access to the substrate. Here we use total internal reflection fluorescence microscopy and nanofabricated DNA curtains to visualize the behavior of Saccharomyces cerevisiae RPA on individual strands of ssDNA in real-time. Our results show that RPA remains bound to ssDNA for long periods of time when free protein is absent from solution. In contrast, RPA rapidly dissociates from ssDNA when free RPA is present in solution allowing rapid exchange between the free and bound states. In addition, the S. cerevisiae DNA recombinase Rad51 and E. coli single-stranded binding protein (SSB) also promote removal of RPA from ssDNA. These results reveal an unanticipated exchange between bound and free RPA suggesting a binding mechanism that can confer exceptionally slow off rates, yet also enables rapid displacement through a direct exchange mechanism that is reliant upon the presence of free ssDNA-binding proteins in solution. Our results indicate that RPA undergoes constant microscopic dissociation under all conditions, but this is only manifested as macroscopic dissociation (i.e. exchange) when free proteins are present in solution, and this effect is due to mass action. We propose that the dissociation of RPA from ssDNA involves a partially dissociated intermediate, which exposes a small section of ssDNA allowing other proteins to access to the DNA.
Conflict of interest statement
Figures
References
-
- Broderick S, Rehmet K, Concannon C, Nasheuer HP (2010) Eukaryotic single-stranded DNA binding proteins: central factors in genome stability. Subcell Biochem 50: 143–163. - PubMed
-
- Wold MS (1997) Replication protein A: a heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu Rev Biochem 66: 61–92. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
