

The role of sleep in emotional brain function
- PMID: 24499013
- PMCID: PMC4286245
- DOI: 10.1146/annurev-clinpsy-032813-153716
The role of sleep in emotional brain function
Abstract
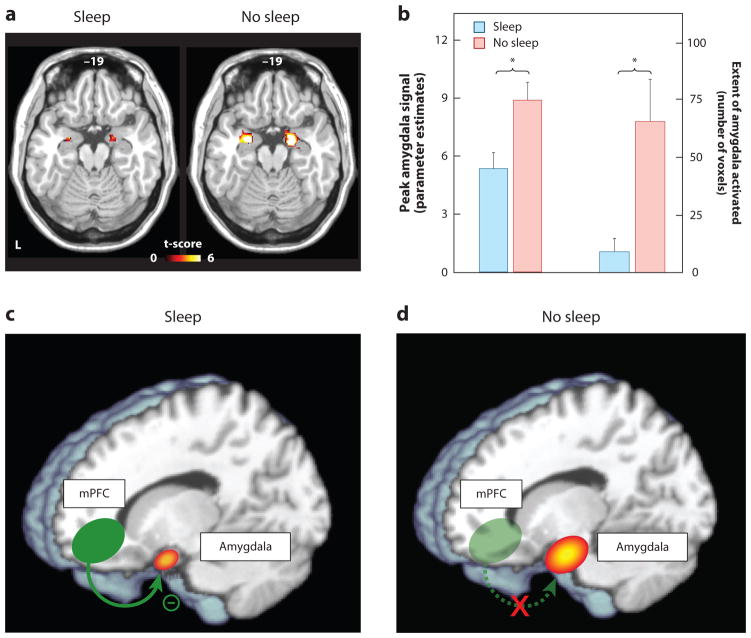
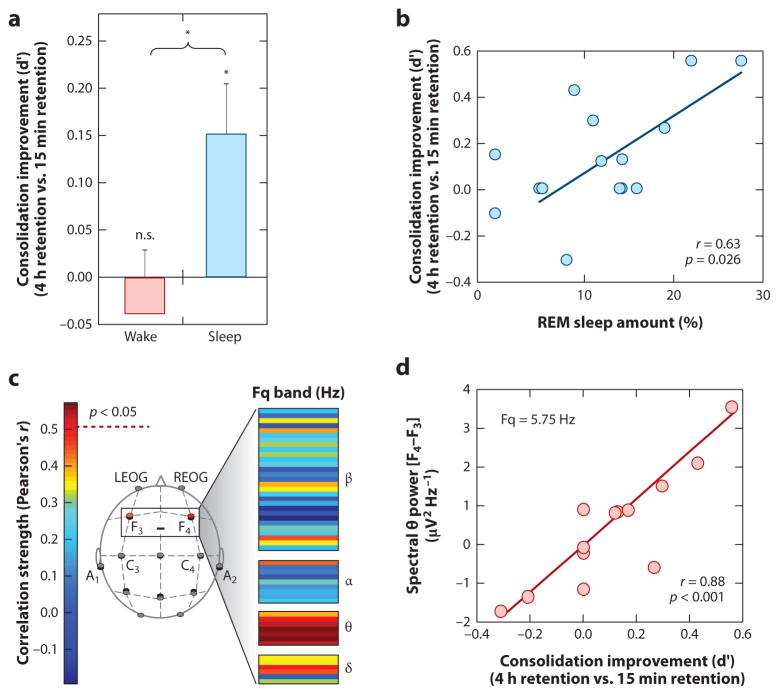
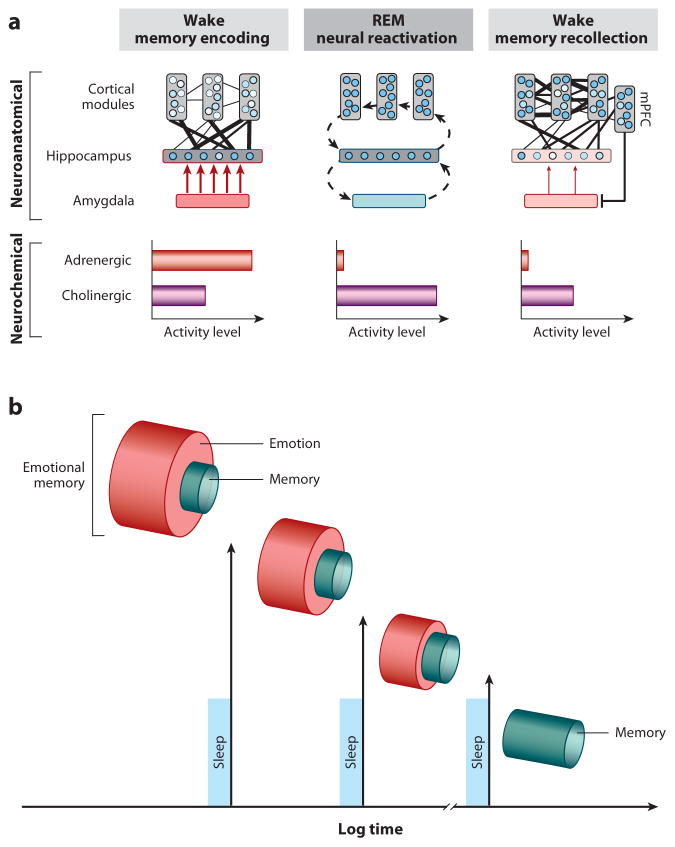
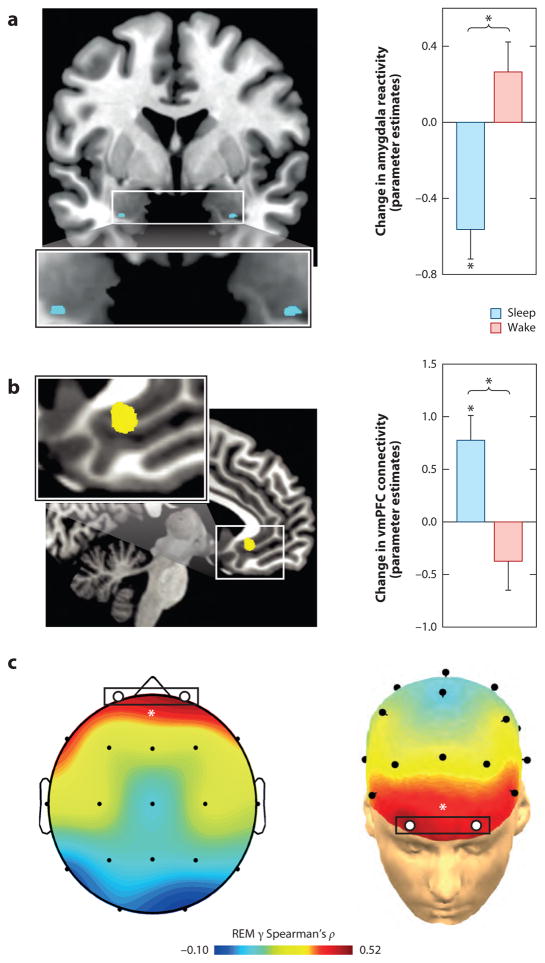
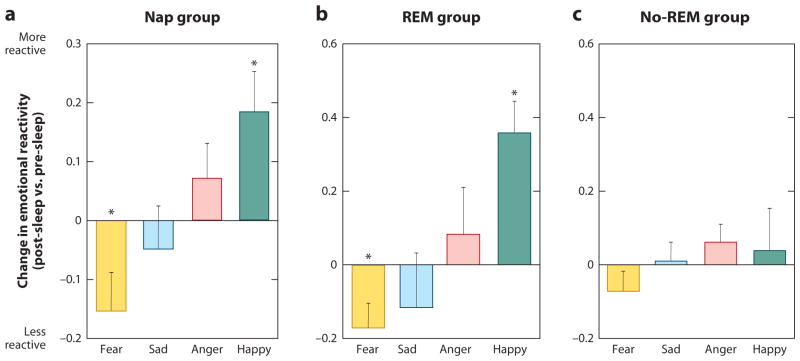
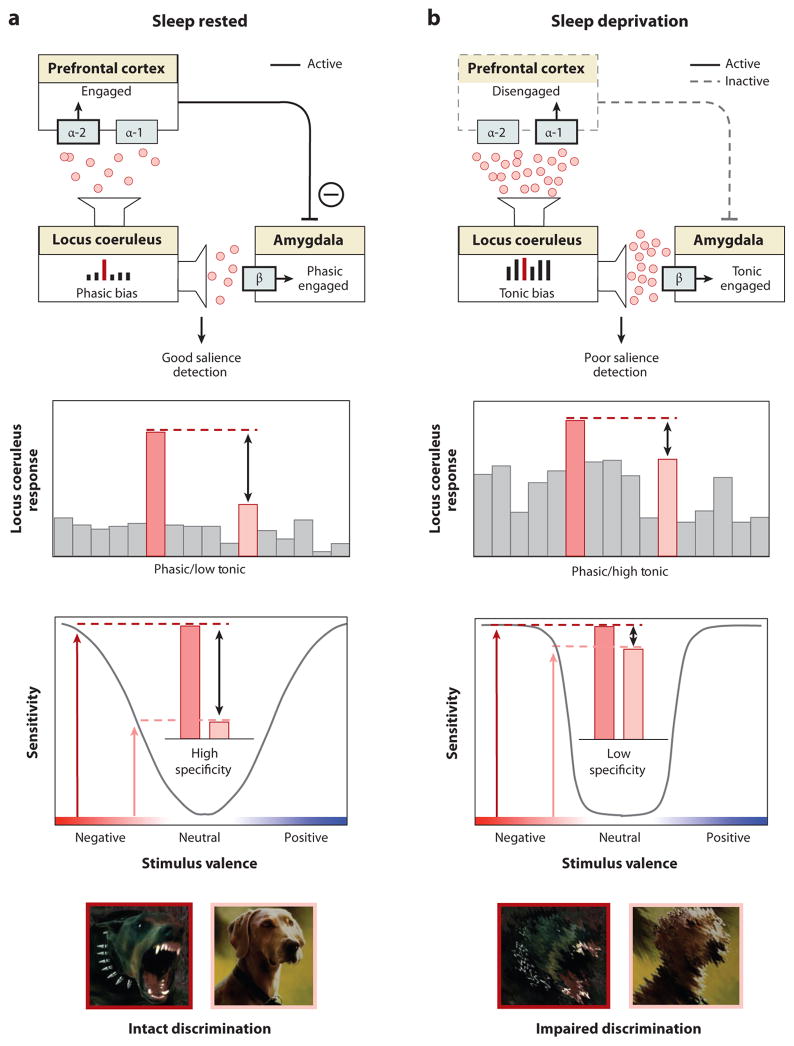
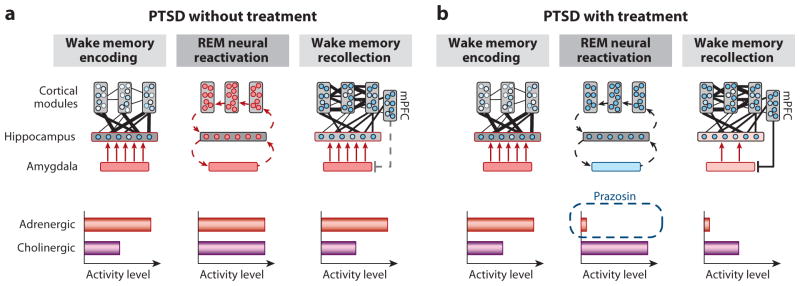
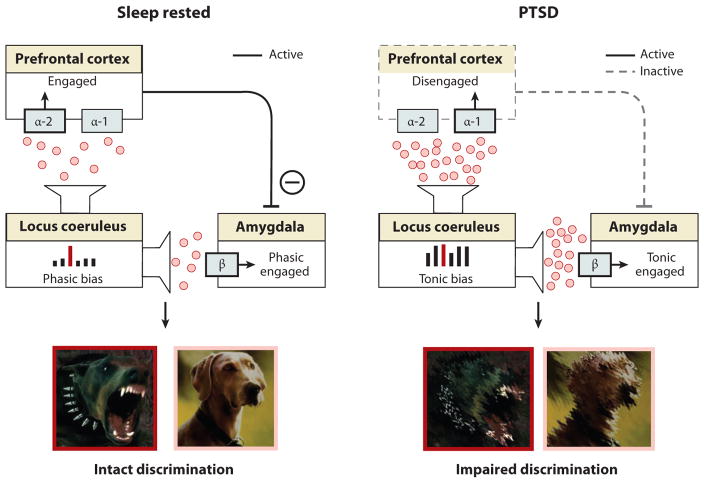
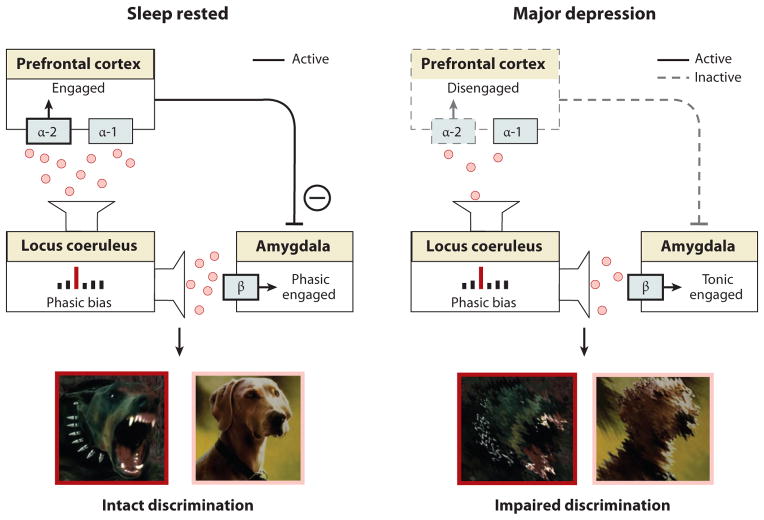
Rapidly emerging evidence continues to describe an intimate and causal relationship between sleep and emotional brain function. These findings are mirrored by long-standing clinical observations demonstrating that nearly all mood and anxiety disorders co-occur with one or more sleep abnormalities. This review aims to (a) provide a synthesis of recent findings describing the emotional brain and behavioral benefits triggered by sleep, and conversely, the detrimental impairments following a lack of sleep; (b) outline a proposed framework in which sleep, and specifically rapid-eye movement (REM) sleep, supports a process of affective brain homeostasis, optimally preparing the organism for next-day social and emotional functioning; and (c) describe how this hypothesized framework can explain the prevalent relationships between sleep and psychiatric disorders, with a particular focus on posttraumatic stress disorder and major depression.
Figures
References
-
- Anderson C, Platten CR. Sleep deprivation lowers inhibition and enhances impulsivity to negative stimuli. Behav Brain Res. 2011;217:463–6. - PubMed
-
- Appelhans B, Luecken L. Heart rate variability as an index of regulated emotional responding. Review of General Psychology. 2006;10:229–40.
-
- Armitage R. Sleep and circadian rhythms in mood disorders. Acta Psychiatr Scand Suppl. 2007:104–15. - PubMed
-
- Aston-Jones G, Cohen JD. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annu Rev Neurosci. 2005;28:403–50. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
