

Proof of principle for epitope-focused vaccine design
- PMID: 24499818
- PMCID: PMC4260937
- DOI: 10.1038/nature12966
Proof of principle for epitope-focused vaccine design
Abstract
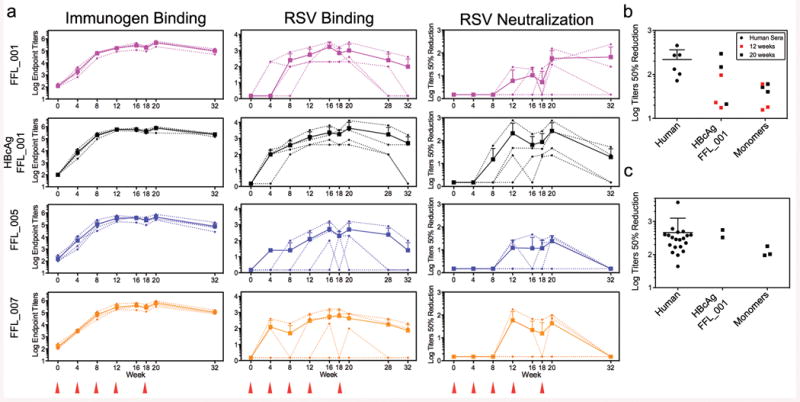
Vaccines prevent infectious disease largely by inducing protective neutralizing antibodies against vulnerable epitopes. Several major pathogens have resisted traditional vaccine development, although vulnerable epitopes targeted by neutralizing antibodies have been identified for several such cases. Hence, new vaccine design methods to induce epitope-specific neutralizing antibodies are needed. Here we show, with a neutralization epitope from respiratory syncytial virus, that computational protein design can generate small, thermally and conformationally stable protein scaffolds that accurately mimic the viral epitope structure and induce potent neutralizing antibodies. These scaffolds represent promising leads for the research and development of a human respiratory syncytial virus vaccine needed to protect infants, young children and the elderly. More generally, the results provide proof of principle for epitope-focused and scaffold-based vaccine design, and encourage the evaluation and further development of these strategies for a variety of other vaccine targets, including antigenically highly variable pathogens such as human immunodeficiency virus and influenza.
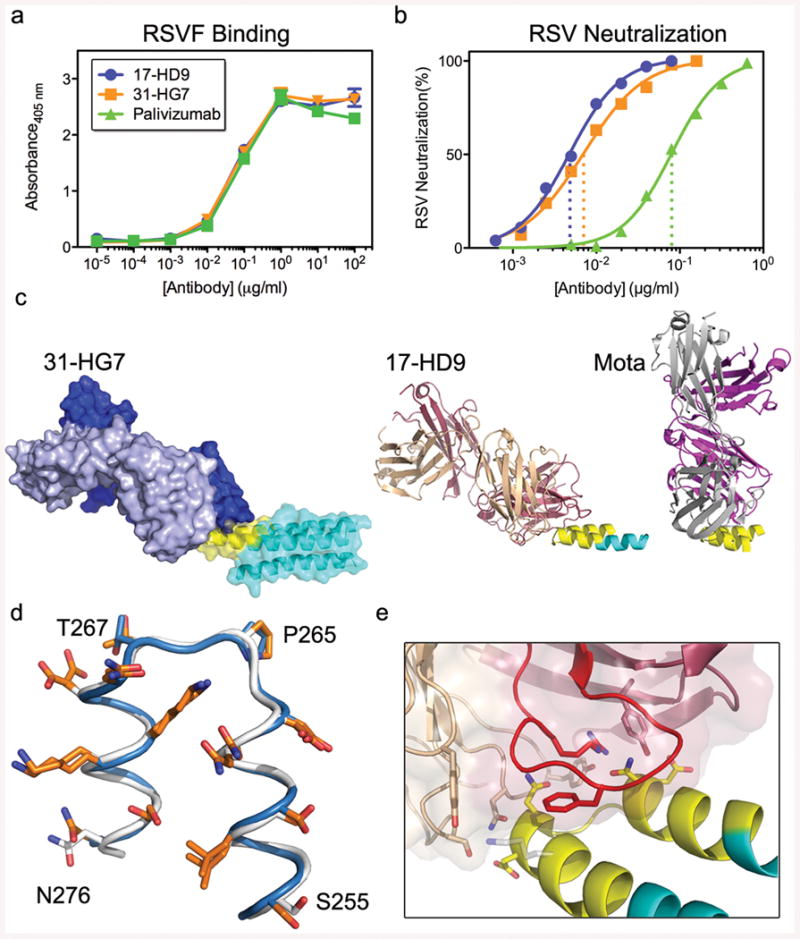
Figures
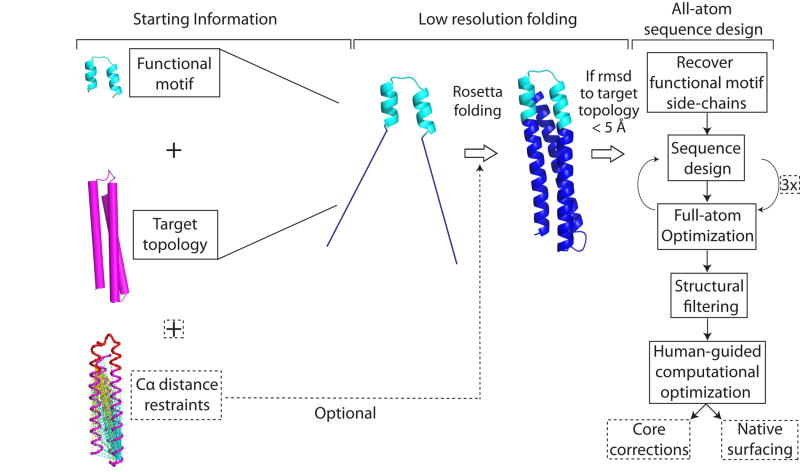
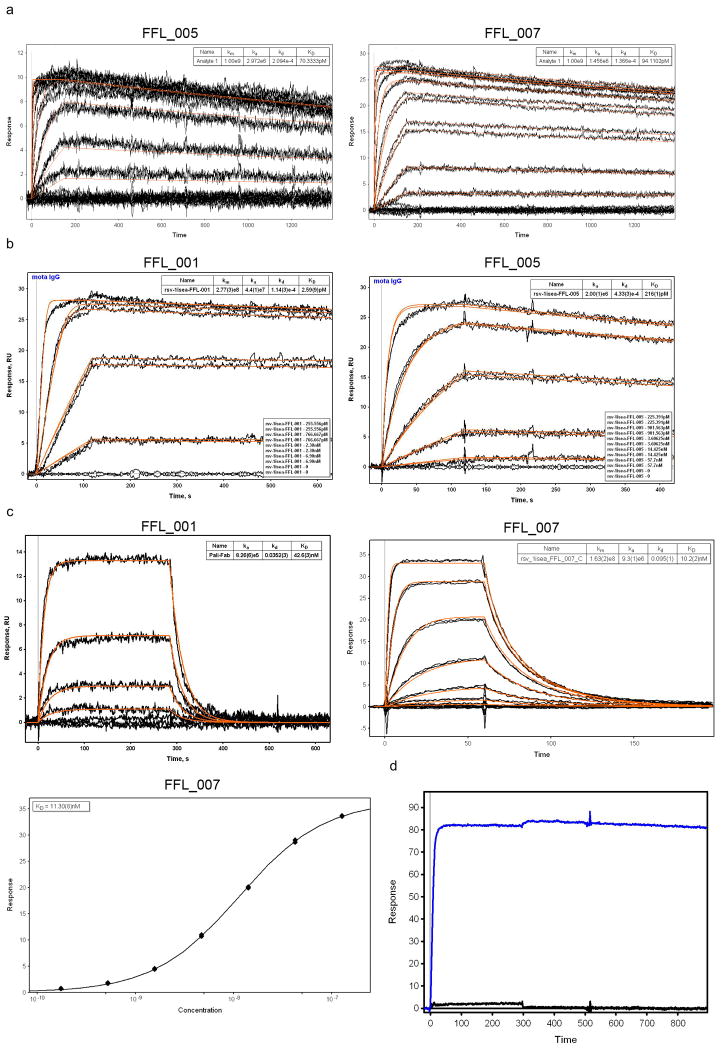

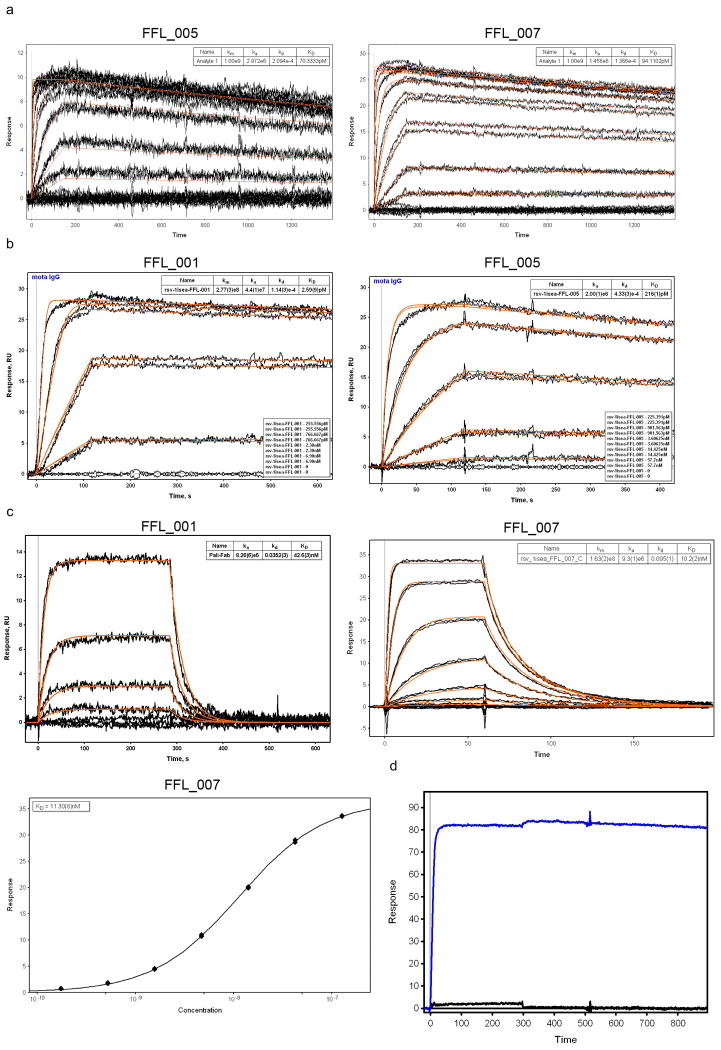
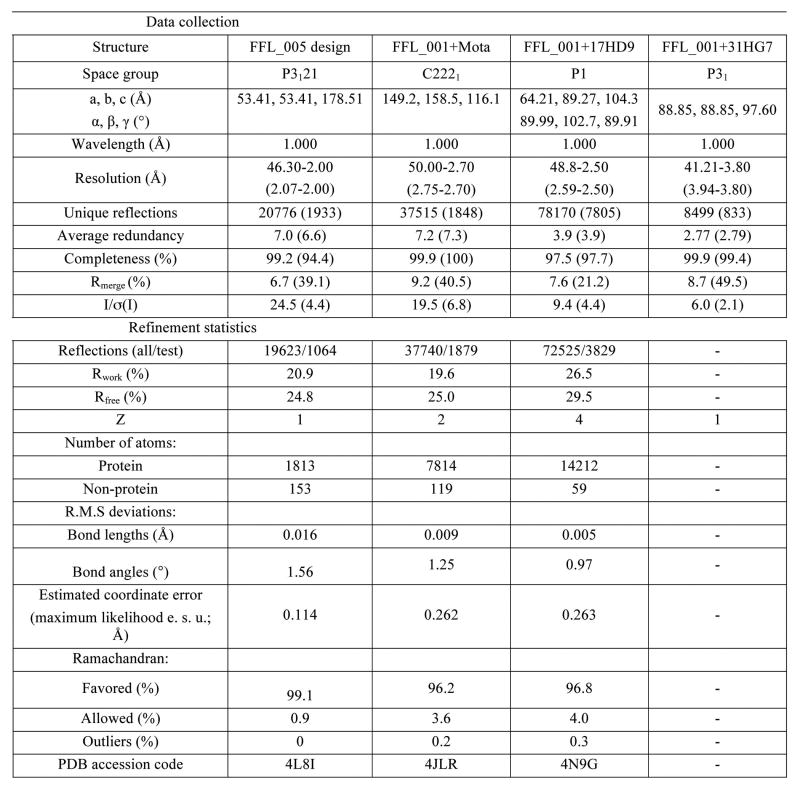
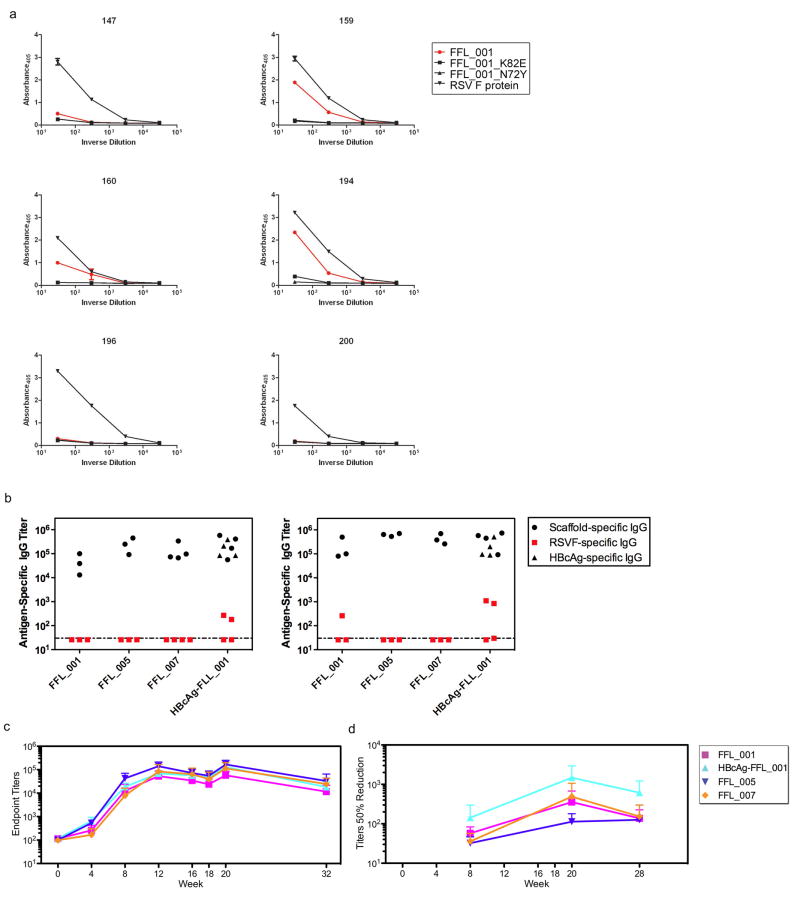
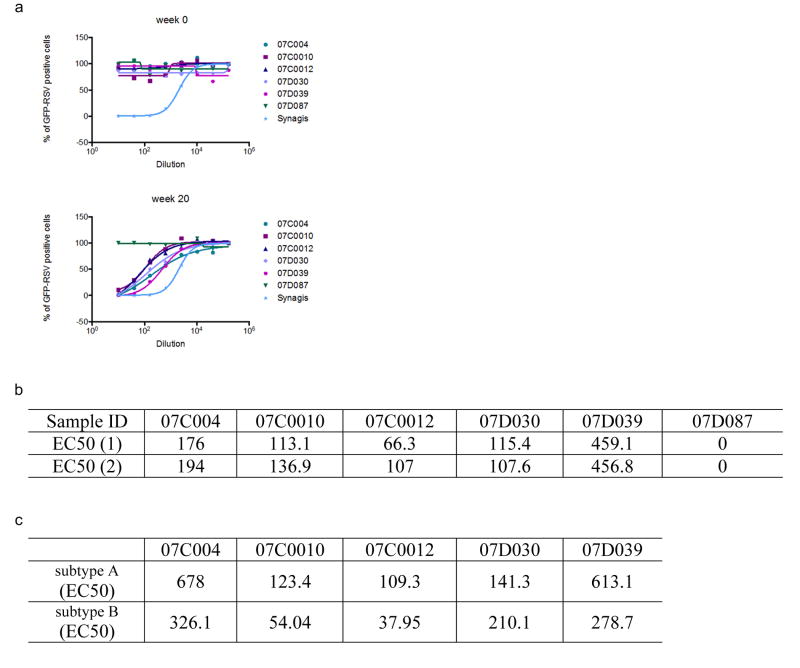
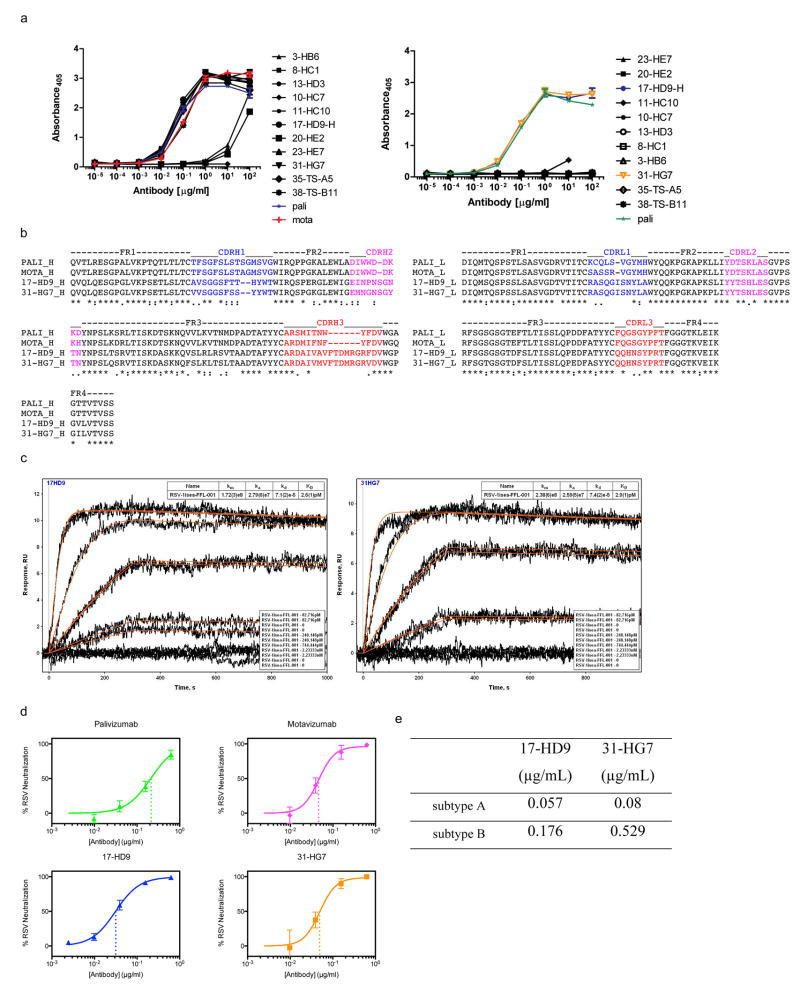
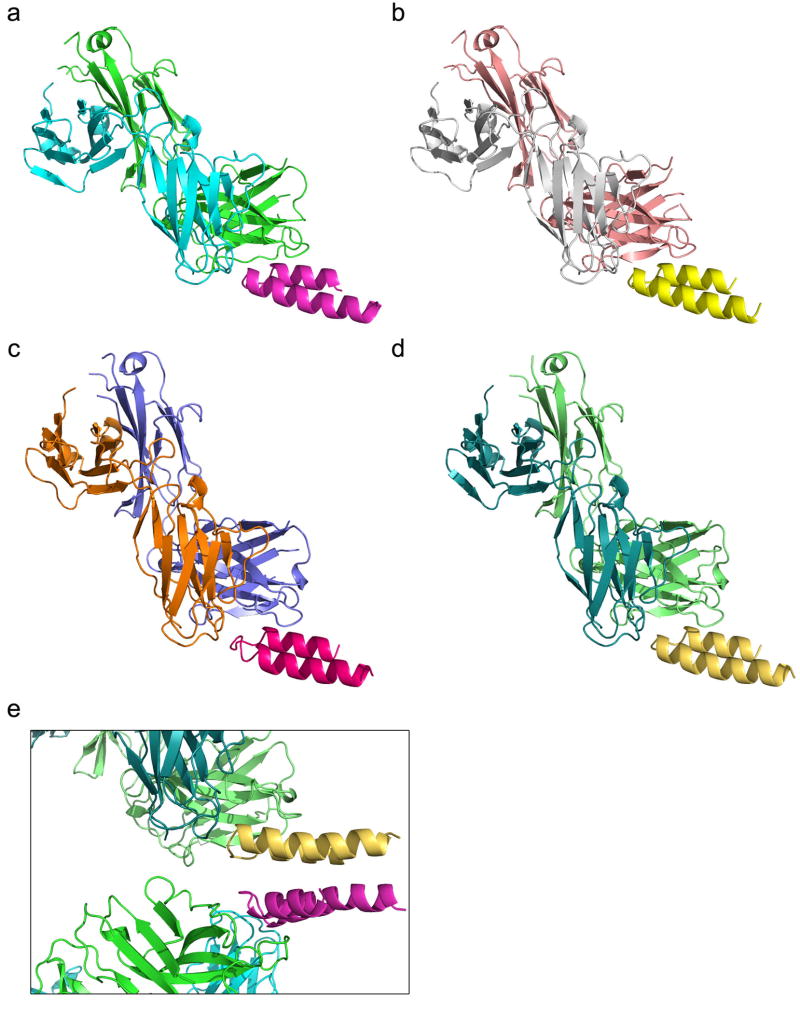
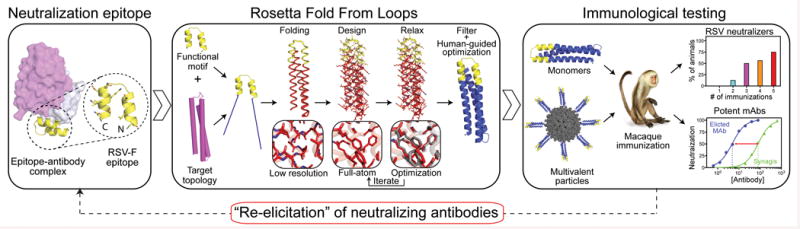
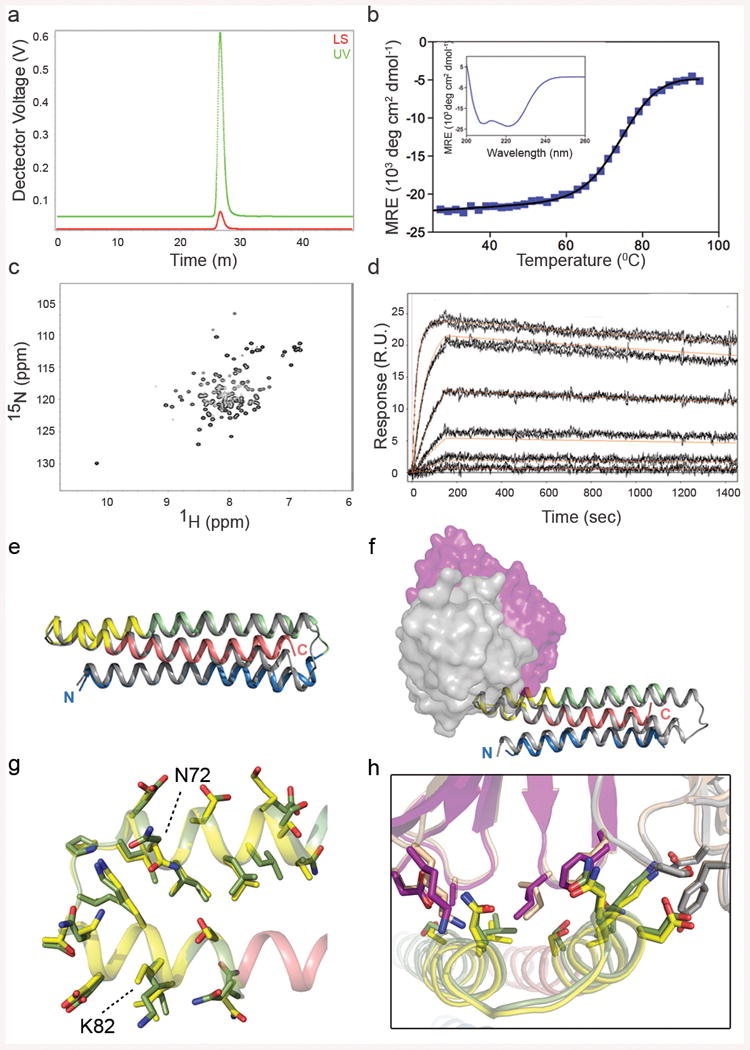
References
-
- Plotkin SA, Orenstein WA, Offit PA. Vaccines. 6. Elsevier; 2012.
-
- Murray CJ, et al. Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990-2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet. 2012;380:2197–2223. - PubMed
-
- Rerks-Ngarm S, et al. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N Engl J Med. 2009;361:2209–2220. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
- U54 AI057141/AI/NIAID NIH HHS/United States
- UM1 AI100663/AI/NIAID NIH HHS/United States
- 2T32GM007270/GM/NIGMS NIH HHS/United States
- R21 AI088554/AI/NIAID NIH HHS/United States
- 5R21AI088554/AI/NIAID NIH HHS/United States
- U54 AI 005714/AI/NIAID NIH HHS/United States
- 1UM1AI100663/AI/NIAID NIH HHS/United States
- T32 GM007270/GM/NIGMS NIH HHS/United States
- ImNIH/Intramural NIH HHS/United States
- P30 AI036214/AI/NIAID NIH HHS/United States
- T32CA080416/CA/NCI NIH HHS/United States
- P01AI094419/AI/NIAID NIH HHS/United States
- P30AI36214/AI/NIAID NIH HHS/United States
- R01 AI102766/AI/NIAID NIH HHS/United States
- P30 AI045008/AI/NIAID NIH HHS/United States
- T32 CA080416/CA/NCI NIH HHS/United States
- 1R01AI102766-01A1/AI/NIAID NIH HHS/United States
- P01 AI094419/AI/NIAID NIH HHS/United States
