

Orsay virus utilizes ribosomal frameshifting to express a novel protein that is incorporated into virions
- PMID: 24503084
- PMCID: PMC3969245
- DOI: 10.1016/j.virol.2013.12.016
Orsay virus utilizes ribosomal frameshifting to express a novel protein that is incorporated into virions
Abstract
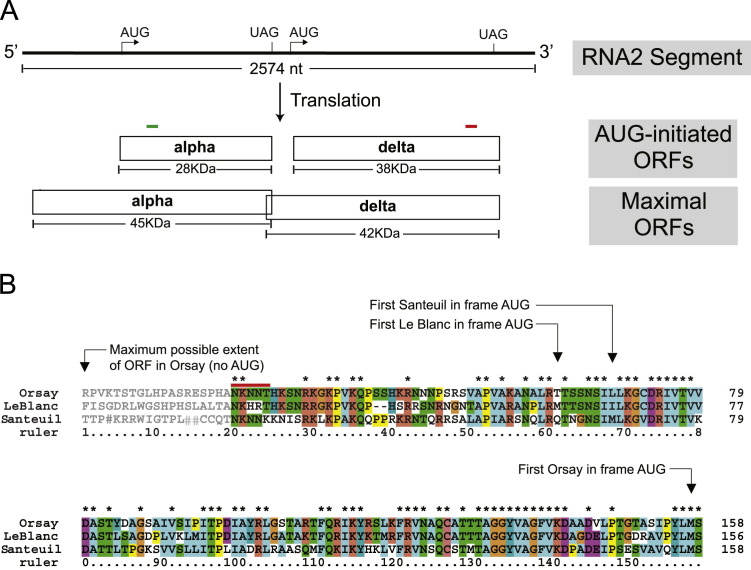
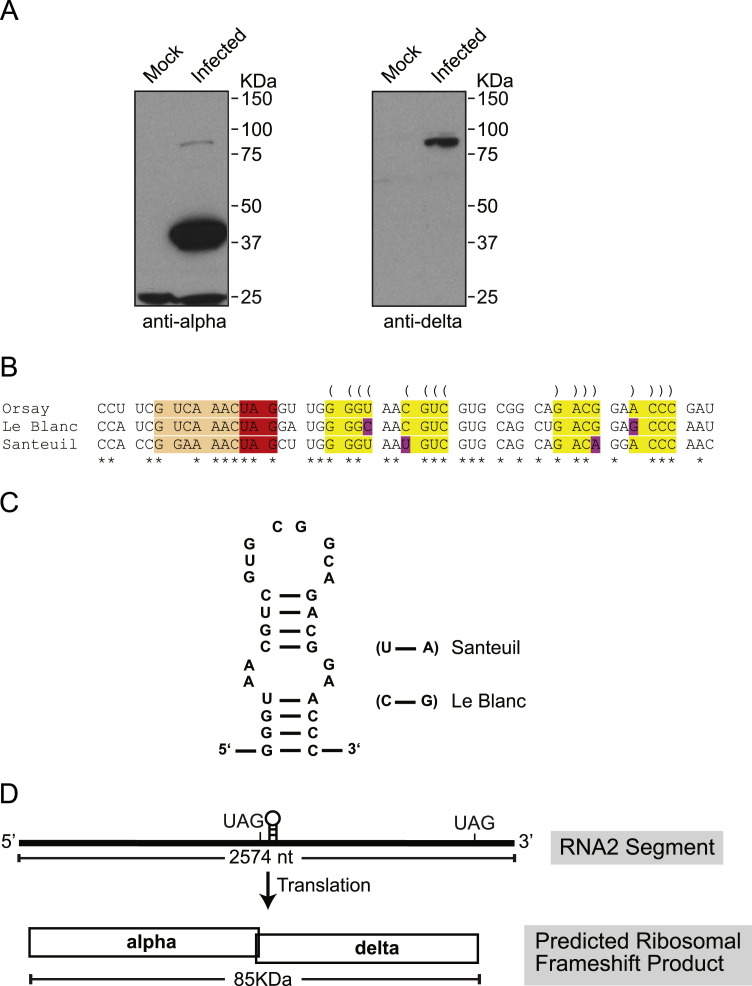
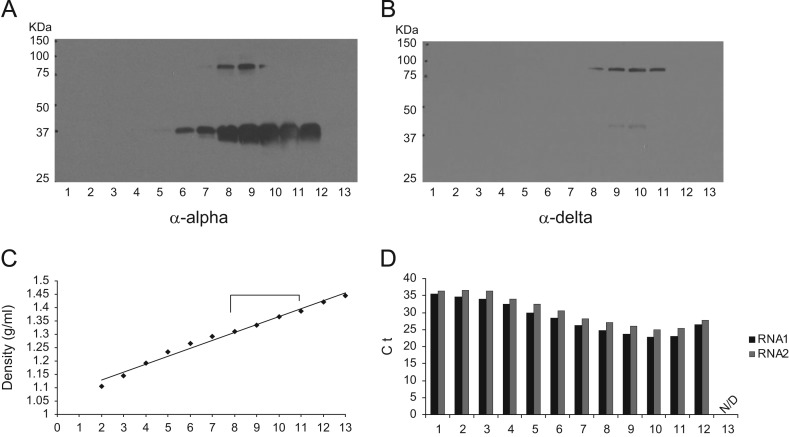
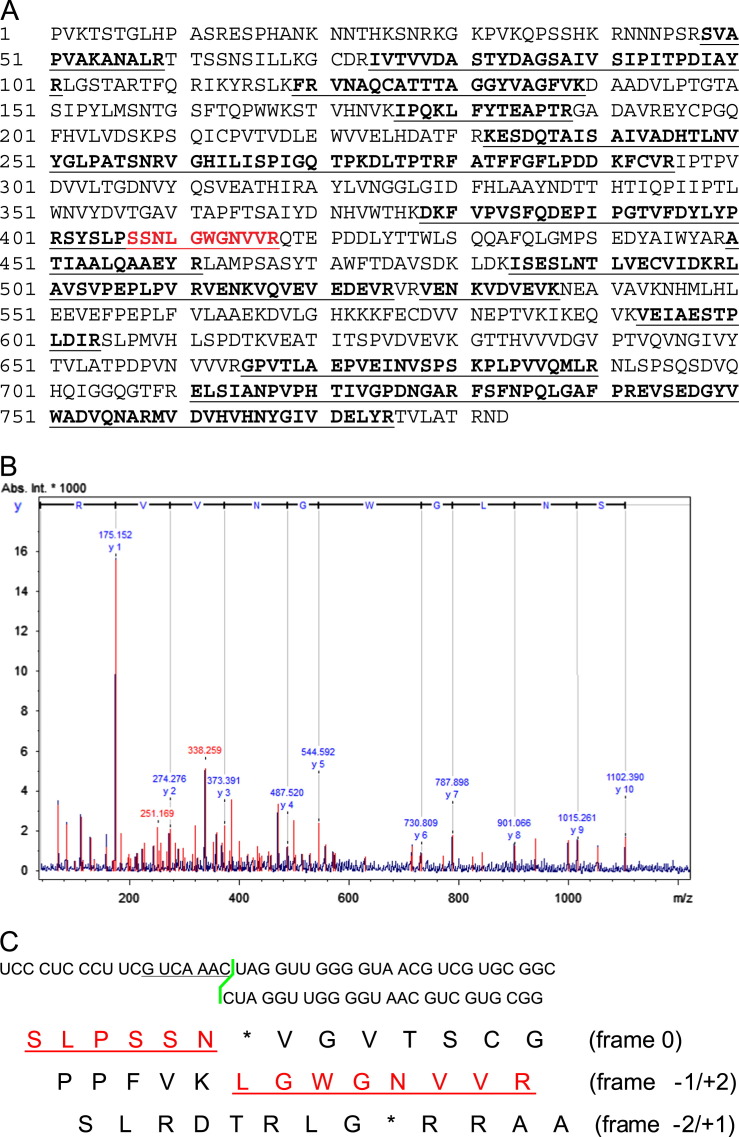
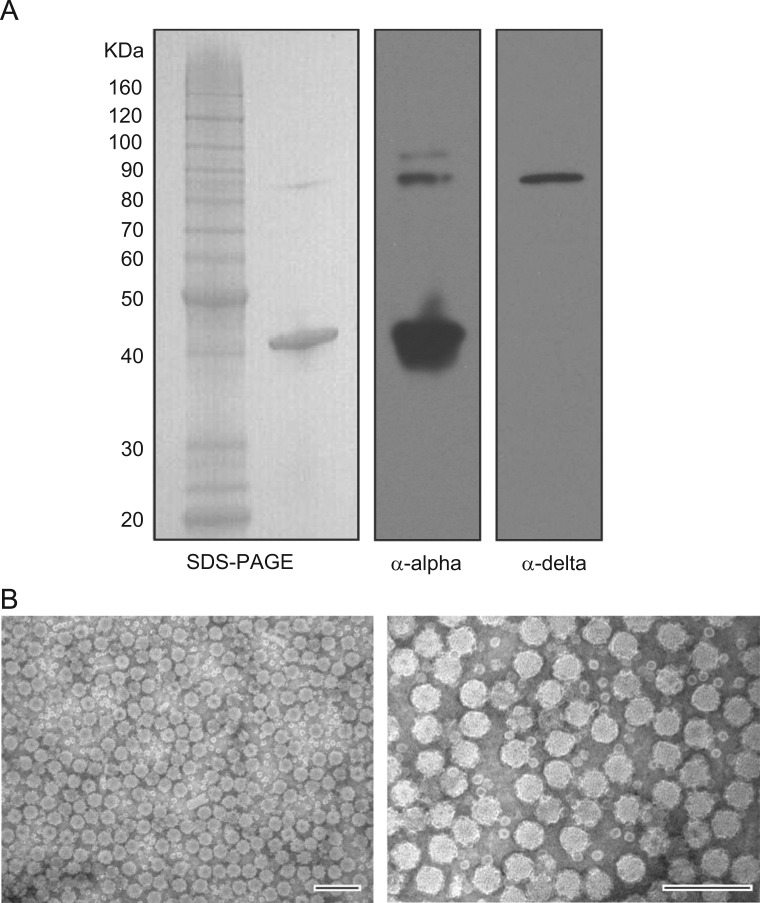
Orsay virus is the first identified virus that is capable of naturally infecting Caenorhabditis elegans. Although it is most closely related to nodaviruses, Orsay virus differs from nodaviruses in its genome organization. In particular, the Orsay virus RNA2 segment encodes a putative novel protein of unknown function, termed delta, which is absent from all known nodaviruses. Here we present evidence that Orsay virus utilizes a ribosomal frameshifting strategy to express a novel fusion protein from the viral capsid (alpha) and delta ORFs. Moreover, the fusion protein was detected in purified virus fractions, demonstrating that it is most likely incorporated into Orsay virions. Furthermore, N-terminal sequencing of both the fusion protein and the capsid protein demonstrated that these proteins must be translated from a non-canonical initiation site. While the function of the alpha-delta fusion remains cryptic, these studies provide novel insights into the fundamental properties of this new clade of viruses.
Keywords: Caenorhabditis elegans; Mass spectrometry; Orsay virus; Ribosomal frameshifting.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Ball L.A., Johnson K.L. Reverse genetics of nodaviruses. Adv. Virus Res. 1999;53:229–244. - PubMed
-
- Brierley I. Ribosomal frameshifting viral RNAs. J. Gen. Virol. 1995;76:1885–1892. - PubMed
-
- Chao J.A., Lee J.H., Chapados B.R., Debler E.W., Schneemann A., Williamson J.R. Dual modes of RNA-silencing suppression by Flock house virus protein B2. Nat. Struct. Mol. Biol. 2005;12:952–957. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
