

Rapid uptake of glucose and lactate, and not hypoxia, induces apoptosis in three-dimensional tumor tissue culture
- PMID: 24503640
- PMCID: PMC4090142
- DOI: 10.1039/c4ib00001c
Rapid uptake of glucose and lactate, and not hypoxia, induces apoptosis in three-dimensional tumor tissue culture
Abstract
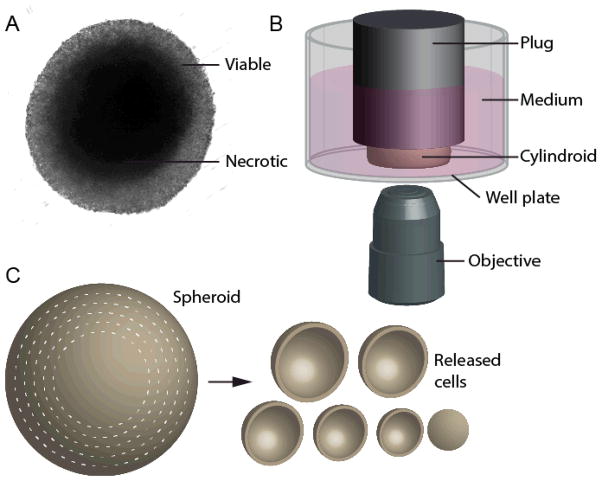
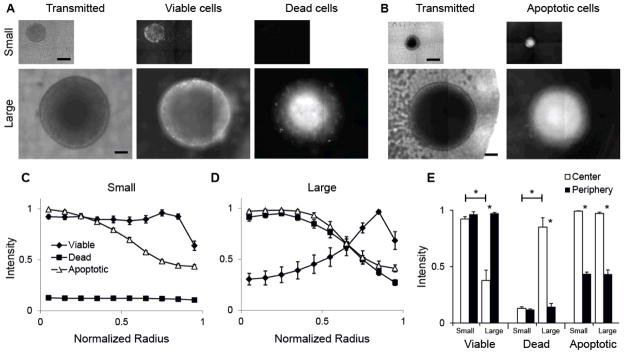
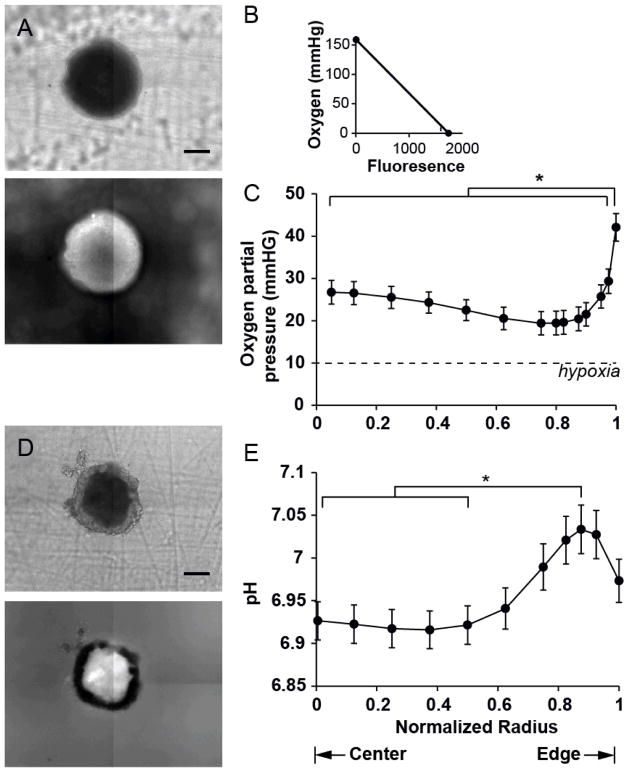
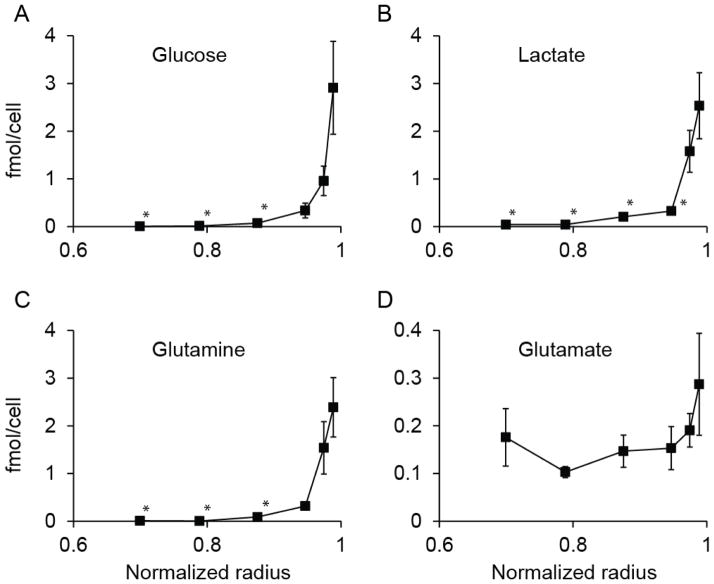
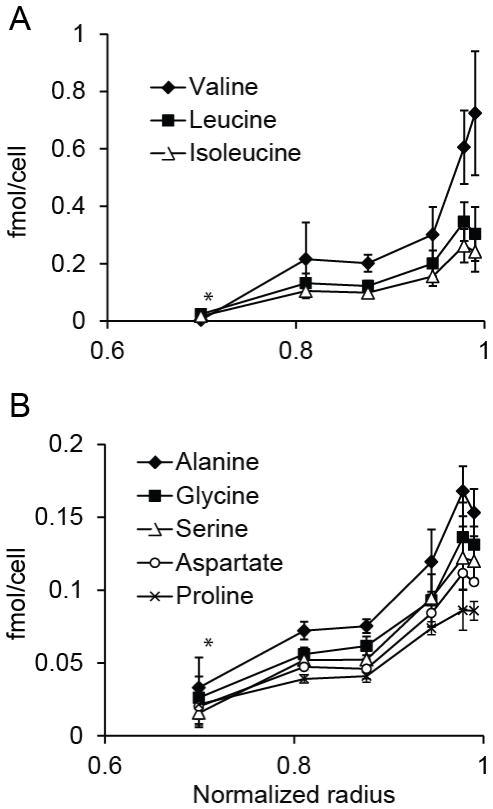
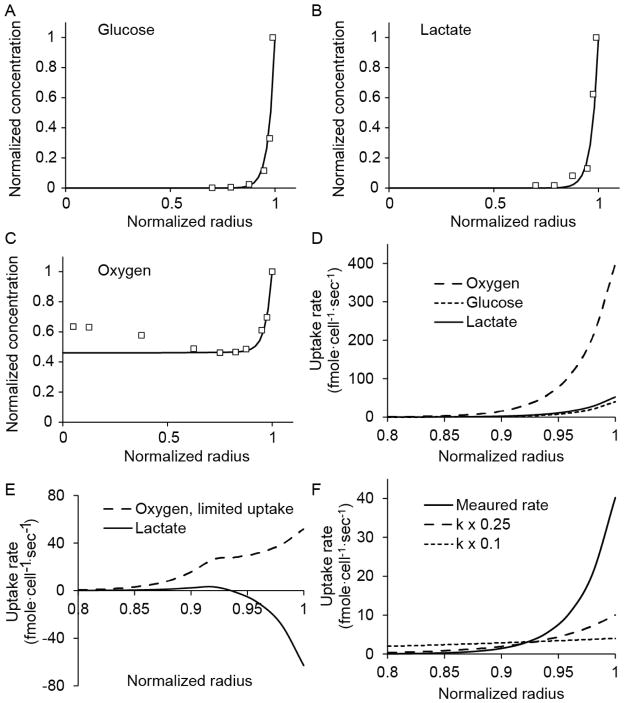
The spatial arrangement of cellular metabolism in tumor tissue critically affects the treatment of cancer. However, little is known about how diffusion and cellular uptake relate to intracellular metabolism and cell death in three dimensions. To quantify these mechanisms, fluorescent microscopy and multicellular tumor cylindroids were used to measure pH and oxygen profiles, and quantify the distribution of viable, apoptotic and necrotic cells. Spheroid dissociation, enzymatic analysis, and mass spectrometry were used to measure concentration profiles of glucose, lactate and glutamine. A mathematical model was used to integrate these measurements and calculate metabolic rate parameters. It was found that large cylindroids, >500 μm in diameter, contained apoptotic and necrotic cells, whereas small cylindroids contained apoptotic but not necrotic cells. The center of cylindroids was found to be acidic but not hypoxic. From the edge to the center, concentrations of glucose, lactate and glutamine decreased rapidly. Throughout the cell masses lactate was consumed and not produced. These measurements indicate that apoptosis was the primary mechanism of cell death; acidity was not caused by lactic acid; and cell death was caused by depletion of carbon sources and not hypoxia. The mathematical model showed that the transporter enzymes for glucose and lactate were not saturated; oxygen uptake was limited by intracellular metabolism; and oxygen uptake was not limited by membrane-transport or diffusion. Unsaturated transmembrane uptake may be the cause of both proliferative and apoptotic regimes in cancer. These results suggest that transporter enzymes are excellent targets for treating well oxygenated tumors.
Figures
Similar articles
-
Evaluating the Metabolic Impact of Hypoxia on Pancreatic Cancer Cells.Methods Mol Biol. 2018;1742:81-93. doi: 10.1007/978-1-4939-7665-2_8. Methods Mol Biol. 2018. PMID: 29330792
-
Single-cell analysis demonstrates how nutrient deprivation creates apoptotic and quiescent cell populations in tumor cylindroids.Biotechnol Bioeng. 2008 Nov 1;101(4):797-810. doi: 10.1002/bit.21985. Biotechnol Bioeng. 2008. PMID: 18814293 Free PMC article.
-
Lactate promotes glutamine uptake and metabolism in oxidative cancer cells.Cell Cycle. 2016;15(1):72-83. doi: 10.1080/15384101.2015.1120930. Cell Cycle. 2016. PMID: 26636483 Free PMC article.
-
Endothelial cell metabolism and tumour angiogenesis: glucose and glutamine as essential fuels and lactate as the driving force.J Intern Med. 2013 Feb;273(2):156-65. doi: 10.1111/joim.12016. J Intern Med. 2013. PMID: 23216817 Review.
-
Hypoxia Dictates Metabolic Rewiring of Tumors: Implications for Chemoresistance.Cells. 2020 Dec 4;9(12):2598. doi: 10.3390/cells9122598. Cells. 2020. PMID: 33291643 Free PMC article. Review.
Cited by
-
Comparing in vitro cytotoxic drug sensitivity in colon and pancreatic cancer using 2D and 3D cell models: Contrasting viability and growth inhibition in clinically relevant dose and repeated drug cycles.Cancer Med. 2024 Jun;13(11):e7318. doi: 10.1002/cam4.7318. Cancer Med. 2024. PMID: 38872378 Free PMC article.
-
Metabolome-Driven Regulation of Adenovirus-Induced Cell Death.Int J Mol Sci. 2021 Jan 5;22(1):464. doi: 10.3390/ijms22010464. Int J Mol Sci. 2021. PMID: 33466472 Free PMC article. Review.
-
Size-Dependent Cortical Compaction Induces Metabolic Adaptation in Mesenchymal Stem Cell Aggregates.Tissue Eng Part A. 2019 Apr;25(7-8):575-587. doi: 10.1089/ten.TEA.2018.0155. Tissue Eng Part A. 2019. PMID: 30187829 Free PMC article.
-
High-Content Monitoring of Drug Effects in a 3D Spheroid Model.Front Oncol. 2017 Dec 11;7:293. doi: 10.3389/fonc.2017.00293. eCollection 2017. Front Oncol. 2017. PMID: 29322028 Free PMC article.
-
A microfluidic device to study cancer metastasis under chronic and intermittent hypoxia.Biomicrofluidics. 2014 Oct 17;8(5):054117. doi: 10.1063/1.4898788. eCollection 2014 Sep. Biomicrofluidics. 2014. PMID: 25584114 Free PMC article.
References
-
- Venkatasubramanian R, Henson MA, Forbes NS. Integrating cell-cycle progression, drug penetration and energy metabolism to identify improved cancer therapeutic strategies. Journal of theoretical biology. 2008;253:98–117. doi: 10.1016/j.jtbi.2008.02.016. - DOI - PMC - PubMed
- Cairns R, Papandreou I, Denko N. Overcoming physiologic barriers to cancer treatment by molecularly targeting the tumor microenvironment. Mol Cancer Res. 2006;4:61–70. - PubMed
-
- Warburg O. Ist die aerobe Glykolyse spezifisch für die Turmoren? Biochem Zietschrift. 1929;204:482.
- Forbes NS, Clark DS, Blanch HW. In: Bioreaction Engineering: Modeling and Control. Schügerl K, Bellgardt KH, editors. Springer Verlag; Berlin: 2000. p Chap 4.4.
-
- Tredan O, Galmarini CM, Patel K, Tannock IF. Drug resistance and the solid tumor microenvironment. J Natl Cancer Inst. 2007;99:1441–54. - PubMed
-
- Cairns RA, Harris IS, Mak TW. Regulation of cancer cell metabolism. Nature Reviews Cancer. 2011;11:85–95. doi: 10.1038/nrc2981. - DOI - PubMed
- Christofk HR, Vander Heiden MG, Harris MH, Ramanathan A, Gerszten RE, Wei R, Fleming MD, Schreiber SL, Cantley LC. The M2 splice isoform of pyruvate kinase is important for cancer metabolism and tumour growth. Nature. 2008;452:230–U74. doi: 10.1038/nature06734. - DOI - PubMed
- Ovens MJ, Davies AJ, Wilson MC, Murray CM, Halestrap AP. AR-C155858 is a potent inhibitor of monocarboxylate transporters MCT1 and MCT2 that binds to an intracellular site involving transmembrane helices 7–10. Biochem J. 2010;425:523–530. doi: 10.1042/bj20091515. - DOI - PMC - PubMed
- Nicklin P, Bergman P, Zhang BL, Triantafellow E, Wang H, Nyfeler B, Yang HD, Hild M, Kung C, Wilson C, Myer VE, MacKeigan JP, Porter JA, Wang YK, Cantley LC, Finan PM, Murphy LO. Bidirectional Transport of Amino Acids Regulates mTOR and Autophagy. Cell. 2009;136:521–534. doi: 10.1016/j.cell.2008.11.044. - DOI - PMC - PubMed
-
- Tennant DA, Duran RV, Gottlieb E. Targeting metabolic ransformation for cancer therapy. Nature Reviews Cancer. 2010;10:267–277. doi: 10.1038/nrc2817. - DOI - PubMed
- Hanai JI, Doro N, Seth P, Sukhatme VP. ATP citrate lyase knockdown impacts cancer stem cells in vitro. Cell Death Dis. 2013;4:e696. doi: 10.1038/cddis.2013.215. - DOI - PMC - PubMed
- Ye JB, Mancuso A, Tong XM, Ward PS, Fan J, Rabinowitz JD, Thompson CB. Pyruvate kinase M2 promotes de novo serine synthesis to sustain mTORC1 activity and cell proliferation. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:6904–6909. doi: 10.1073/pnas.1204176109. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
