

Uterine selection of human embryos at implantation
- PMID: 24503642
- PMCID: PMC3915549
- DOI: 10.1038/srep03894
Uterine selection of human embryos at implantation
Abstract
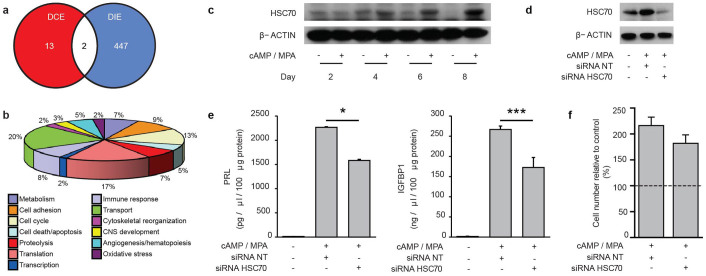
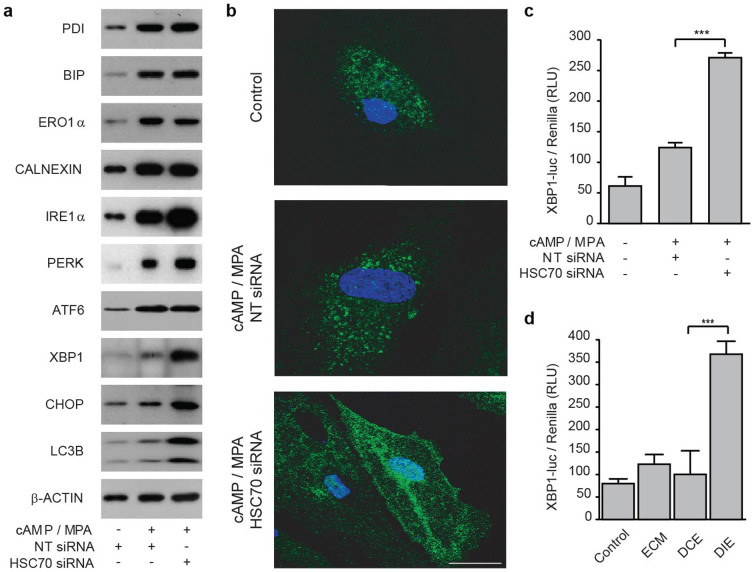
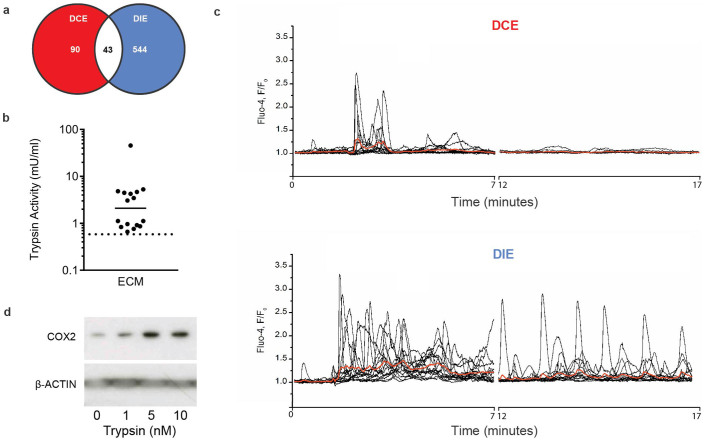
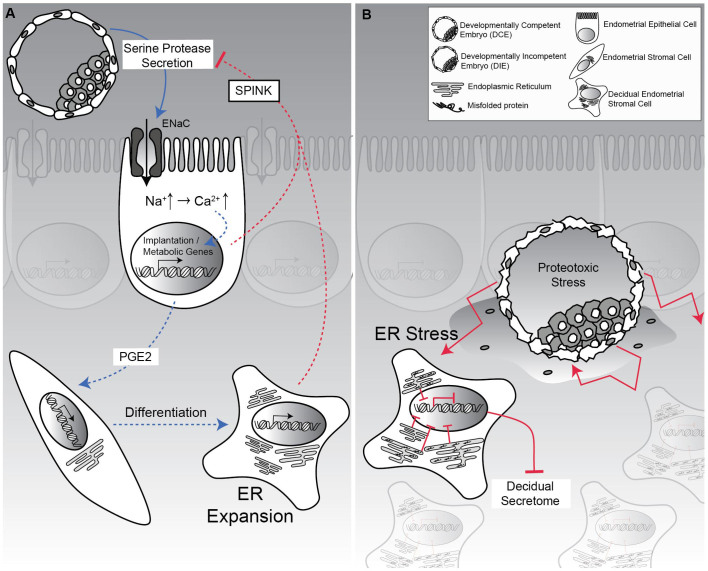
Human embryos frequently harbor large-scale complex chromosomal errors that impede normal development. Affected embryos may fail to implant although many first breach the endometrial epithelium and embed in the decidualizing stroma before being rejected via mechanisms that are poorly understood. Here we show that developmentally impaired human embryos elicit an endoplasmic stress response in human decidual cells. A stress response was also evident upon in vivo exposure of mouse uteri to culture medium conditioned by low-quality human embryos. By contrast, signals emanating from developmentally competent embryos activated a focused gene network enriched in metabolic enzymes and implantation factors. We further show that trypsin, a serine protease released by pre-implantation embryos, elicits Ca(2+) signaling in endometrial epithelial cells. Competent human embryos triggered short-lived oscillatory Ca(2+) fluxes whereas low-quality embryos caused a heightened and prolonged Ca(2+) response. Thus, distinct positive and negative mechanisms contribute to active selection of human embryos at implantation.
Figures
References
-
- Macklon N. S., Geraedts J. P. & Fauser B. C. Conception to ongoing pregnancy: the ‘black box’ of early pregnancy loss. Hum Reprod Update 8, 333–343 (2002). - PubMed
-
- Rai R. & Regan L. Recurrent miscarriage. Lancet 368, 601–611 (2006). - PubMed
-
- Fragouli E. et al. The origin and impact of embryonic aneuploidy. Hum Genet, 10.1007/s00439-013-1309-0 (2013). - PubMed
-
- Mertzanidou A. et al. Microarray analysis reveals abnormal chromosomal complements in over 70% of 14 normally developing human embryos. Hum Reprod 28, 256–264, 10.1093/humrep/des362 (2013). - PubMed
-
- Vanneste E. et al. Chromosome instability is common in human cleavage-stage embryos. Nat Med 15, 577–583 (2009). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
